

Ex-527 inhibits Sirtuins by exploiting their unique NAD+-dependent deacetylation mechanism
- PMID: 23840057
- PMCID: PMC3725051
- DOI: 10.1073/pnas.1303628110
Ex-527 inhibits Sirtuins by exploiting their unique NAD+-dependent deacetylation mechanism
Abstract
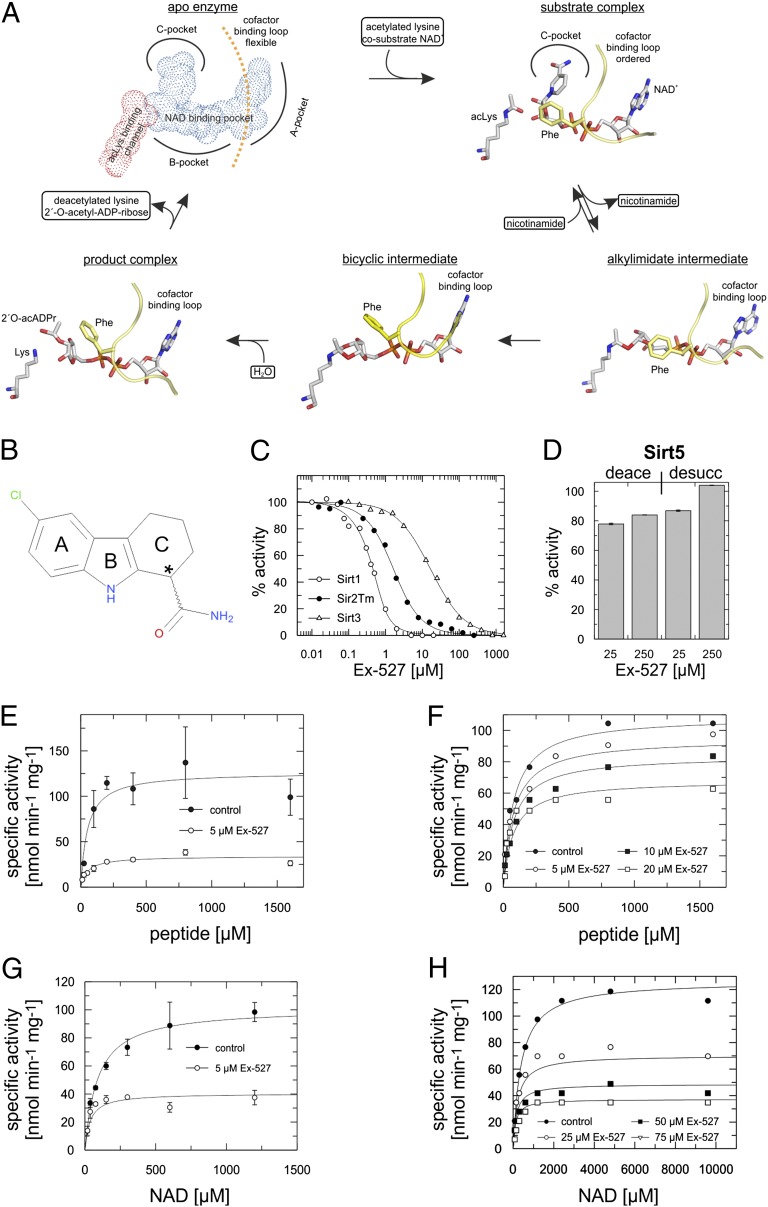
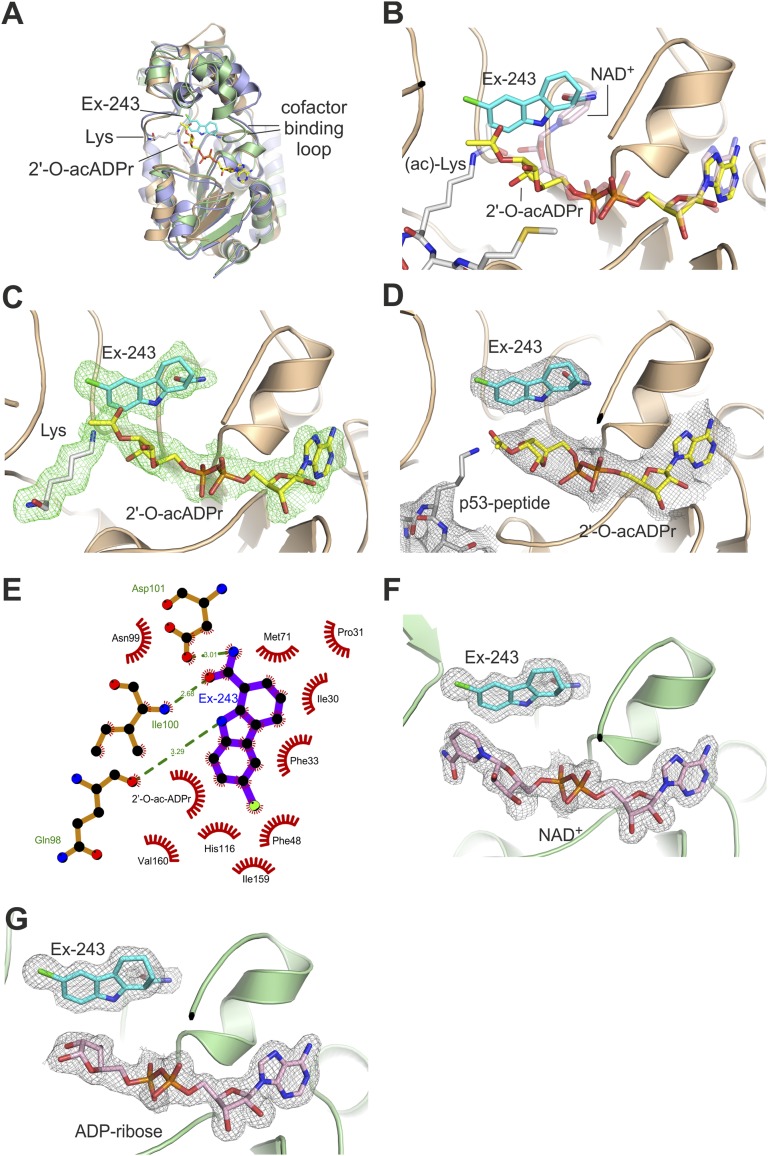
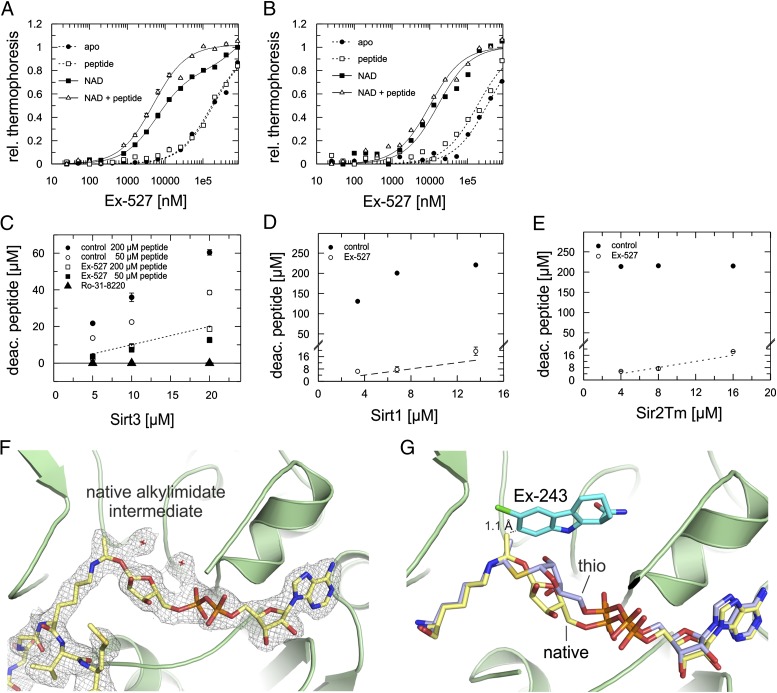
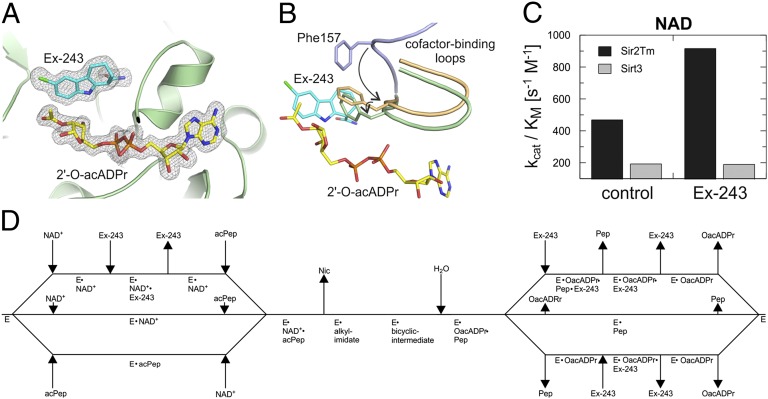
Sirtuins are protein deacetylases regulating metabolism and stress responses. The seven human Sirtuins (Sirt1-7) are attractive drug targets, but Sirtuin inhibition mechanisms are mostly unidentified. We report the molecular mechanism of Sirtuin inhibition by 6-chloro-2,3,4,9-tetrahydro-1H-carbazole-1-carboxamide (Ex-527). Inhibitor binding to potently inhibited Sirt1 and Thermotoga maritima Sir2 and to moderately inhibited Sirt3 requires NAD(+), alone or together with acetylpeptide. Crystal structures of several Sirtuin inhibitor complexes show that Ex-527 occupies the nicotinamide site and a neighboring pocket and contacts the ribose of NAD(+) or of the coproduct 2'-O-acetyl-ADP ribose. Complex structures with native alkylimidate and thio-analog support its catalytic relevance and show, together with biochemical assays, that only the coproduct complex is relevant for inhibition by Ex-527, which stabilizes the closed enzyme conformation preventing product release. Ex-527 inhibition thus exploits Sirtuin catalysis, and kinetic isoform differences explain its selectivity. Our results provide insights in Sirtuin catalysis and inhibition with important implications for drug development.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Substrate specificity and kinetic mechanism of the Sir2 family of NAD+-dependent histone/protein deacetylases.Biochemistry. 2004 Aug 3;43(30):9877-87. doi: 10.1021/bi049592e. Biochemistry. 2004. PMID: 15274642
-
Kinetic and Structural Basis for Acyl-Group Selectivity and NAD(+) Dependence in Sirtuin-Catalyzed Deacylation.Biochemistry. 2015 May 19;54(19):3037-3050. doi: 10.1021/acs.biochem.5b00150. Epub 2015 May 4. Biochemistry. 2015. PMID: 25897714 Free PMC article.
-
The 2.5 Å crystal structure of the SIRT1 catalytic domain bound to nicotinamide adenine dinucleotide (NAD+) and an indole (EX527 analogue) reveals a novel mechanism of histone deacetylase inhibition.J Med Chem. 2013 Feb 14;56(3):963-9. doi: 10.1021/jm301431y. Epub 2013 Jan 29. J Med Chem. 2013. PMID: 23311358
-
SIR2: the biochemical mechanism of NAD(+)-dependent protein deacetylation and ADP-ribosyl enzyme intermediates.Curr Med Chem. 2004 Apr;11(7):807-26. doi: 10.2174/0929867043455675. Curr Med Chem. 2004. PMID: 15078167 Review.
-
Small molecule regulation of Sir2 protein deacetylases.FEBS J. 2005 Sep;272(18):4607-16. doi: 10.1111/j.1742-4658.2005.04862.x. FEBS J. 2005. PMID: 16156783 Review.
Cited by
-
Sirtuin 1 inhibits TNF-α-mediated osteoclastogenesis of bone marrow-derived macrophages through both ROS generation and TRPV1 activation.Mol Cell Biochem. 2019 May;455(1-2):135-145. doi: 10.1007/s11010-018-3477-7. Epub 2018 Nov 20. Mol Cell Biochem. 2019. PMID: 30456702
-
Differentiated modulation of signaling molecules AMPK and SIRT1 in experimentally drug-induced hepatocyte injury.Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2023 Mar;167(1):50-60. doi: 10.5507/bp.2022.018. Epub 2022 Apr 12. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub. 2023. PMID: 35416184
-
Myricanol rescues dexamethasone-induced muscle dysfunction via a sirtuin 1-dependent mechanism.J Cachexia Sarcopenia Muscle. 2019 Apr;10(2):429-444. doi: 10.1002/jcsm.12393. Epub 2019 Feb 21. J Cachexia Sarcopenia Muscle. 2019. PMID: 30793539 Free PMC article.
-
The roles of sirtuins in ferroptosis.Front Physiol. 2023 Apr 20;14:1131201. doi: 10.3389/fphys.2023.1131201. eCollection 2023. Front Physiol. 2023. PMID: 37153222 Free PMC article. Review.
-
p53 Proteoforms and Intrinsic Disorder: An Illustration of the Protein Structure-Function Continuum Concept.Int J Mol Sci. 2016 Nov 10;17(11):1874. doi: 10.3390/ijms17111874. Int J Mol Sci. 2016. PMID: 27834926 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
