

Coupling unbiased mutagenesis to high-throughput DNA sequencing uncovers functional domains in the Ndc80 kinetochore protein of Saccharomyces cerevisiae
- PMID: 23833183
- PMCID: PMC3761298
- DOI: 10.1534/genetics.113.152728
Coupling unbiased mutagenesis to high-throughput DNA sequencing uncovers functional domains in the Ndc80 kinetochore protein of Saccharomyces cerevisiae
Abstract
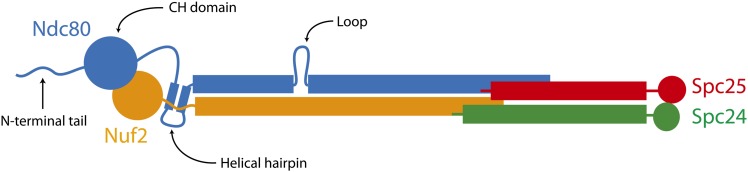
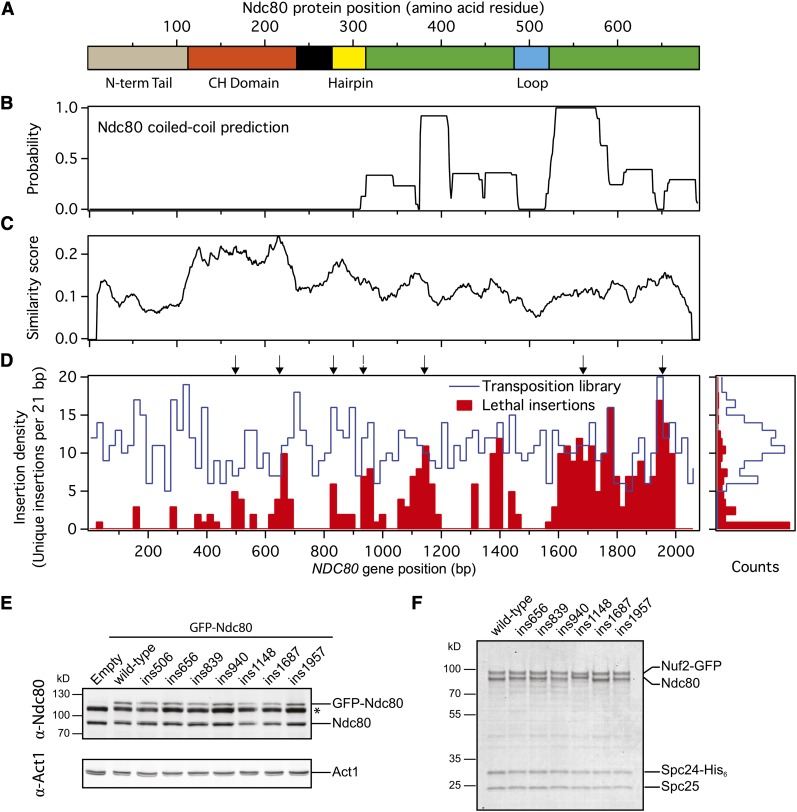
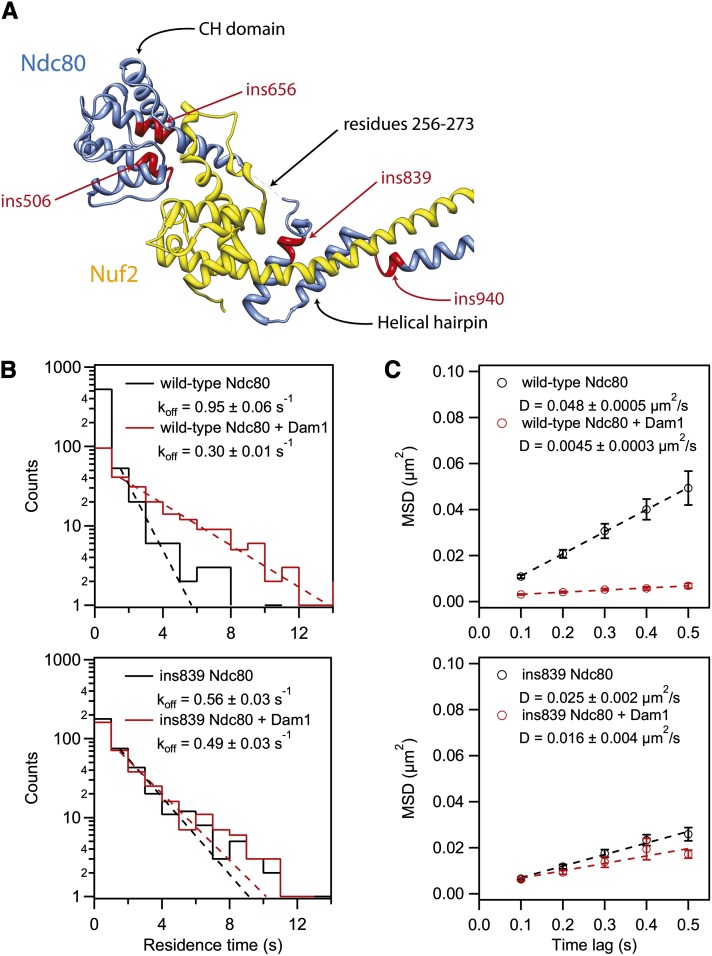
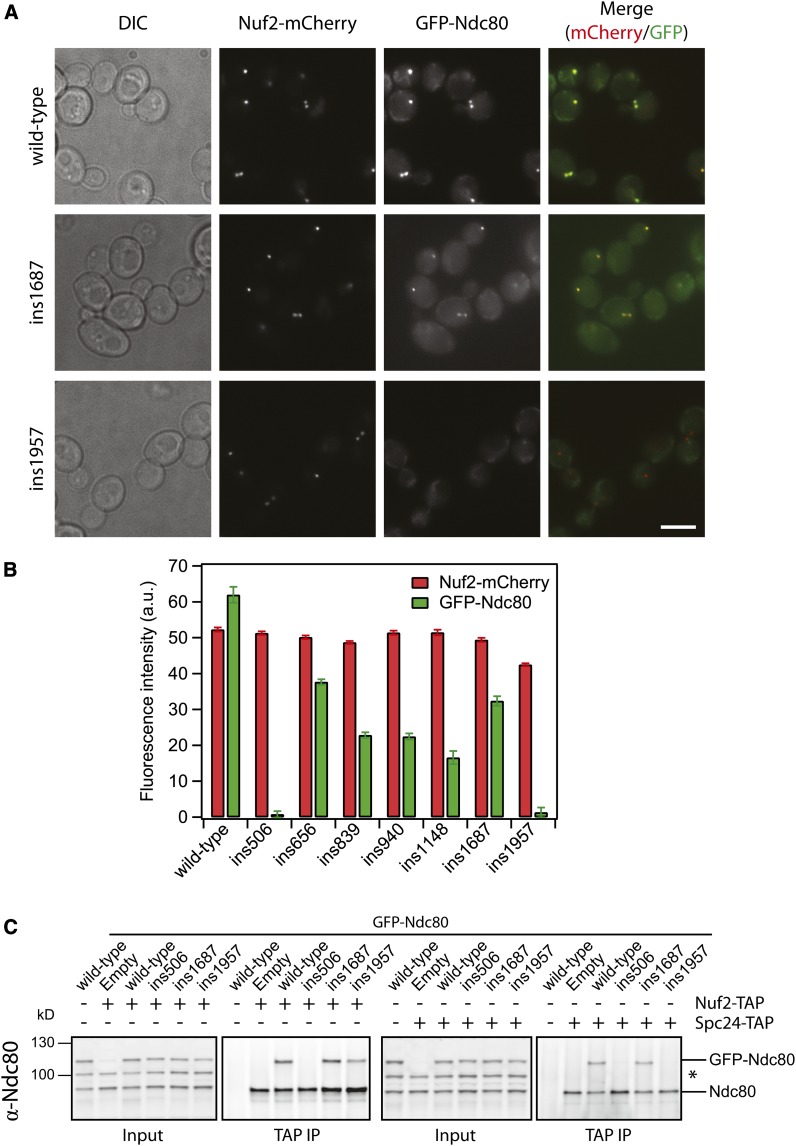
During mitosis, kinetochores physically link chromosomes to the dynamic ends of spindle microtubules. This linkage depends on the Ndc80 complex, a conserved and essential microtubule-binding component of the kinetochore. As a member of the complex, the Ndc80 protein forms microtubule attachments through a calponin homology domain. Ndc80 is also required for recruiting other components to the kinetochore and responding to mitotic regulatory signals. While the calponin homology domain has been the focus of biochemical and structural characterization, the function of the remainder of Ndc80 is poorly understood. Here, we utilized a new approach that couples high-throughput sequencing to a saturating linker-scanning mutagenesis screen in Saccharomyces cerevisiae. We identified domains in previously uncharacterized regions of Ndc80 that are essential for its function in vivo. We show that a helical hairpin adjacent to the calponin homology domain influences microtubule binding by the complex. Furthermore, a mutation in this hairpin abolishes the ability of the Dam1 complex to strengthen microtubule attachments made by the Ndc80 complex. Finally, we defined a C-terminal segment of Ndc80 required for tetramerization of the Ndc80 complex in vivo. This unbiased mutagenesis approach can be generally applied to genes in S. cerevisiae to identify functional properties and domains.
Keywords: Hec1; Illumina; coiled coil; total internal reflection fluorescence (TIRF).
Figures

Similar articles
-
Kinetochore biorientation in Saccharomyces cerevisiae requires a tightly folded conformation of the Ndc80 complex.Genetics. 2014 Dec;198(4):1483-93. doi: 10.1534/genetics.114.167775. Epub 2014 Sep 16. Genetics. 2014. PMID: 25230952 Free PMC article.
-
Molecular requirements for the formation of a kinetochore-microtubule interface by Dam1 and Ndc80 complexes.J Cell Biol. 2013 Jan 7;200(1):21-30. doi: 10.1083/jcb.201210091. Epub 2012 Dec 31. J Cell Biol. 2013. PMID: 23277429 Free PMC article.
-
Cooperation of the Dam1 and Ndc80 kinetochore complexes enhances microtubule coupling and is regulated by aurora B.J Cell Biol. 2010 May 17;189(4):713-23. doi: 10.1083/jcb.200910142. J Cell Biol. 2010. PMID: 20479468 Free PMC article.
-
The Ndc80 complex: hub of kinetochore activity.FEBS Lett. 2007 Jun 19;581(15):2862-9. doi: 10.1016/j.febslet.2007.05.012. Epub 2007 May 11. FEBS Lett. 2007. PMID: 17521635 Review.
-
Structural view of the yeast Dam1 complex, a ring-shaped molecular coupler for the dynamic microtubule end.Essays Biochem. 2020 Sep 4;64(2):359-370. doi: 10.1042/EBC20190079. Essays Biochem. 2020. PMID: 32579171 Free PMC article. Review.
Cited by
-
Tight bending of the Ndc80 complex provides intrinsic regulation of its binding to microtubules.Elife. 2019 May 2;8:e44489. doi: 10.7554/eLife.44489. Elife. 2019. PMID: 31045495 Free PMC article.
-
Three interacting regions of the Ndc80 and Dam1 complexes support microtubule tip-coupling under load.J Cell Biol. 2022 May 2;221(5):e202107016. doi: 10.1083/jcb.202107016. Epub 2022 Mar 30. J Cell Biol. 2022. PMID: 35353161 Free PMC article.
-
Measuring mitotic forces.Methods Cell Biol. 2018;144:165-184. doi: 10.1016/bs.mcb.2018.03.007. Epub 2018 Apr 10. Methods Cell Biol. 2018. PMID: 29804669 Free PMC article.
-
Structural mechanism of outer kinetochore Dam1-Ndc80 complex assembly on microtubules.Science. 2023 Dec 8;382(6675):1184-1190. doi: 10.1126/science.adj8736. Epub 2023 Dec 7. Science. 2023. PMID: 38060647 Free PMC article.
-
Novel phosphorylation states of the yeast spindle pole body.Biol Open. 2018 Oct 8;7(10):bio033647. doi: 10.1242/bio.033647. Biol Open. 2018. PMID: 29903865 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
