

Lipid synthesis in protozoan parasites: a comparison between kinetoplastids and apicomplexans
- PMID: 23827884
- PMCID: PMC3830643
- DOI: 10.1016/j.plipres.2013.06.003
Lipid synthesis in protozoan parasites: a comparison between kinetoplastids and apicomplexans
Abstract
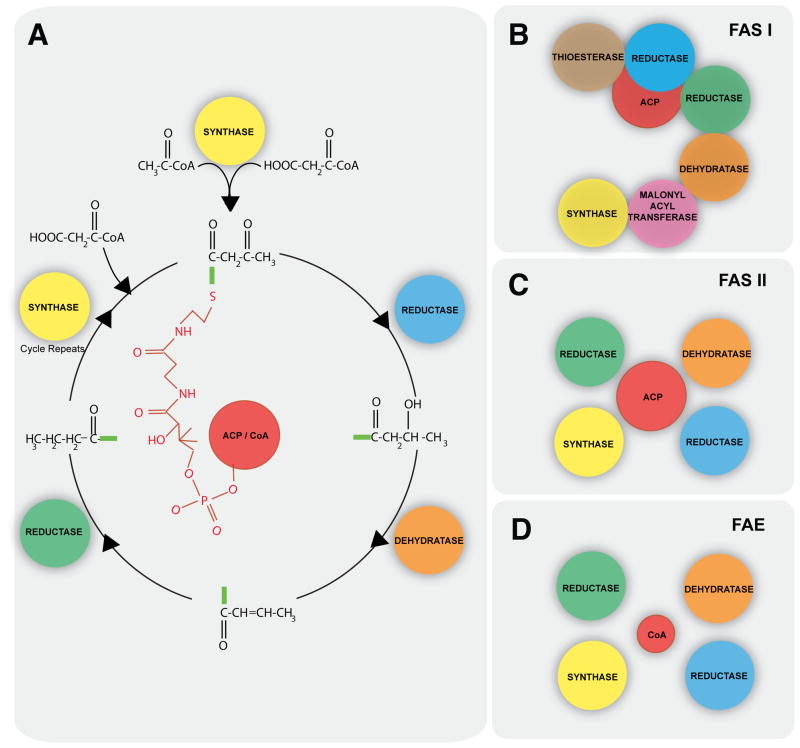
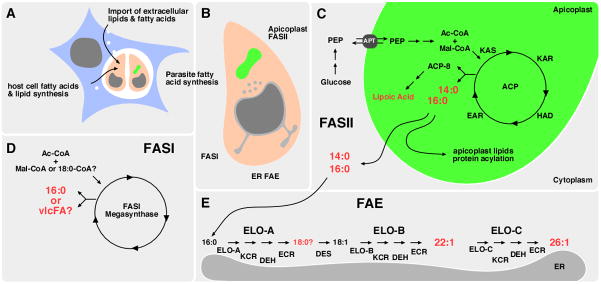
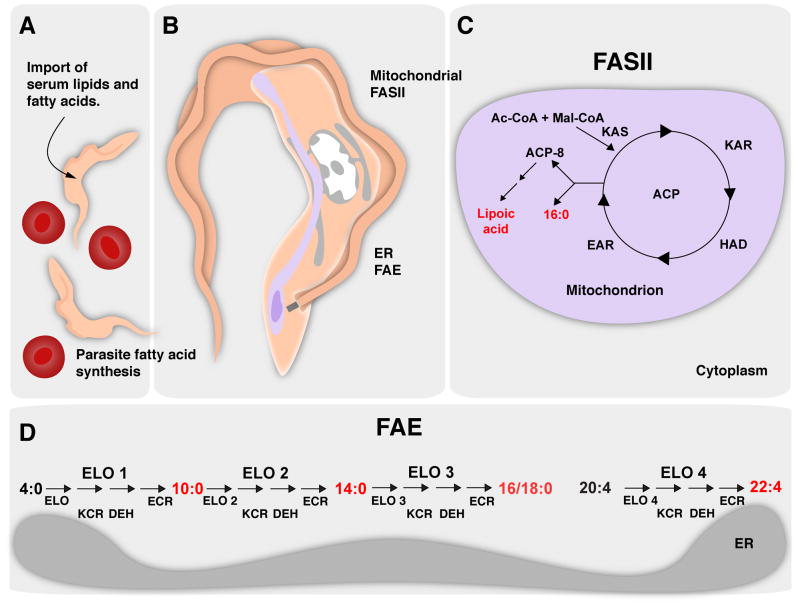
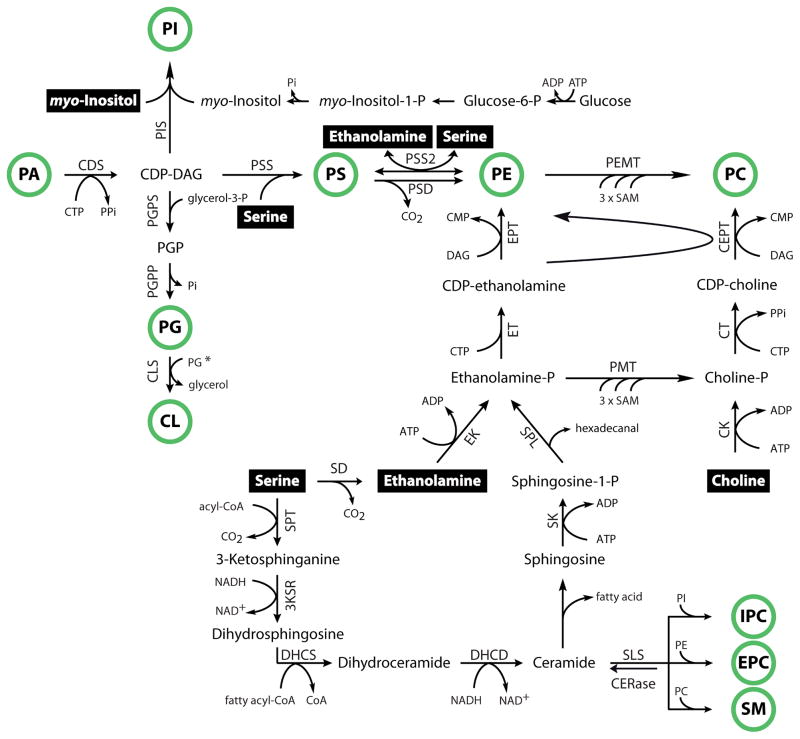
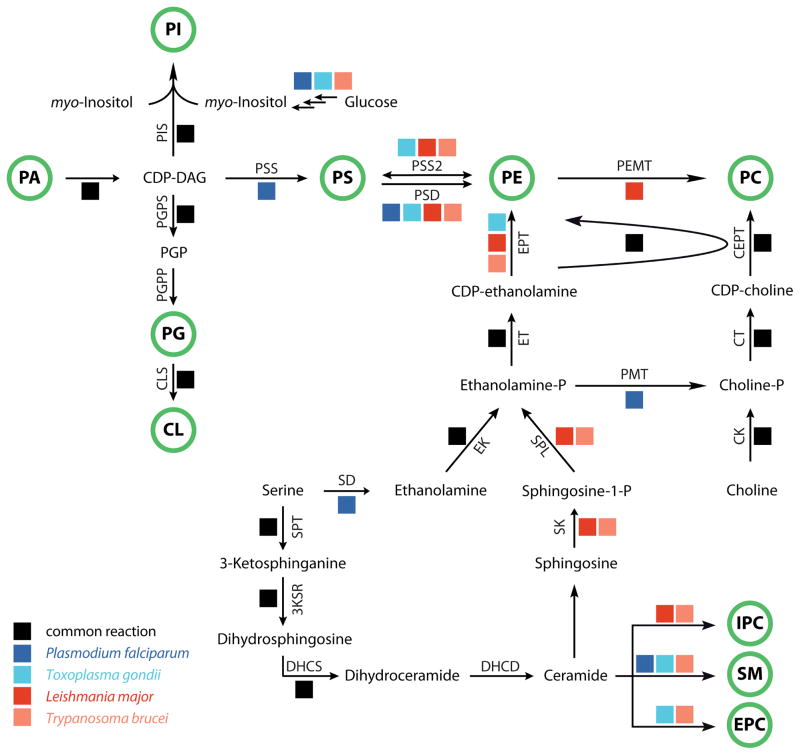
Lipid metabolism is of crucial importance for pathogens. Lipids serve as cellular building blocks, signalling molecules, energy stores, posttranslational modifiers, and pathogenesis factors. Parasites rely on a complex system of uptake and synthesis mechanisms to satisfy their lipid needs. The parameters of this system change dramatically as the parasite transits through the various stages of its life cycle. Here we discuss the tremendous recent advances that have been made in the understanding of the synthesis and uptake pathways for fatty acids and phospholipids in apicomplexan and kinetoplastid parasites, including Plasmodium, Toxoplasma, Cryptosporidium, Trypanosoma and Leishmania. Lipid synthesis differs in significant ways between parasites from both phyla and the human host. Parasites have acquired novel pathways through endosymbiosis, as in the case of the apicoplast, have dramatically reshaped substrate and product profiles, and have evolved specialized lipids to interact with or manipulate the host. These differences potentially provide opportunities for drug development. We outline the lipid pathways for key species in detail as they progress through the developmental cycle and highlight those that are of particular importance to the biology of the pathogens and/or are the most promising targets for parasite-specific treatment.
Keywords: ACC; ACP; Apicomplexa; CDP-choline/ethanolamine:diacylglycerol phosphotransferase; CDP-ethanolamine:diacylglycerol phosphotransferase; CEPT; CL; Drugs; EPC; EPT; ER; FAE; FASI; FASII; Fatty acid synthesis; GPI; IPC; Kinetoplastida; MSP1; MUFAs; PC; PE; PG; PGP; PI; PKS; PS; PUFAs; Phospholipid synthesis; Protozoa; RNA interference; RNAi; SCD; SM; TS; acetyl-CoA carboxylase; acyl carrier protein; cardiolipin; endoplasmic reticulum; ethanolamine phosphorylceramide; fatty acid elongation; fatty acid synthase type I; fatty acid synthase type II; glycosylphosphatidylinositol; inositol phosphorylceramide; merozoite surface protein-1; monounsaturated fatty acids; phosphatidylcholine; phosphatidylethanolamine; phosphatidylglycerol; phosphatidylglycerophosphate; phosphatidylinositol; phosphatidylserine; polyketide synthase; polyunsaturated fatty acids; sphingomyelin; stearoyl-CoA desaturase; thiastearates.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Toxoplasma gondii acyl-lipid metabolism: de novo synthesis from apicoplast-generated fatty acids versus scavenging of host cell precursors.Biochem J. 2006 Feb 15;394(Pt 1):197-205. doi: 10.1042/BJ20050609. Biochem J. 2006. PMID: 16246004 Free PMC article.
-
The role of phospholipids in the biological activity and structure of the endoplasmic reticulum.Biochim Biophys Acta. 2013 Nov;1833(11):2499-510. doi: 10.1016/j.bbamcr.2013.05.018. Epub 2013 May 24. Biochim Biophys Acta. 2013. PMID: 23711956 Review.
-
The acyl-CoA synthetase TgACS1 allows neutral lipid metabolism and extracellular motility in Toxoplasma gondii through relocation via its peroxisomal targeting sequence (PTS) under low nutrient conditions.mBio. 2024 Apr 10;15(4):e0042724. doi: 10.1128/mbio.00427-24. Epub 2024 Mar 19. mBio. 2024. PMID: 38501871 Free PMC article.
-
Current progress in the fatty acid metabolism in Cryptosporidium parvum.J Eukaryot Microbiol. 2004 Jul-Aug;51(4):381-8. doi: 10.1111/j.1550-7408.2004.tb00384.x. J Eukaryot Microbiol. 2004. PMID: 15352319 Review.
-
Apicoplast and endoplasmic reticulum cooperate in fatty acid biosynthesis in apicomplexan parasite Toxoplasma gondii.J Biol Chem. 2012 Feb 10;287(7):4957-71. doi: 10.1074/jbc.M111.310144. Epub 2011 Dec 16. J Biol Chem. 2012. PMID: 22179608 Free PMC article.
Cited by
-
Characterization of metabolically quiescent Leishmania parasites in murine lesions using heavy water labeling.PLoS Pathog. 2015 Feb 25;11(2):e1004683. doi: 10.1371/journal.ppat.1004683. eCollection 2015 Feb. PLoS Pathog. 2015. PMID: 25714830 Free PMC article.
-
Nematode phospholipid metabolism: an example of closing the genome-structure-function circle.Trends Parasitol. 2014 May;30(5):241-50. doi: 10.1016/j.pt.2014.03.001. Epub 2014 Mar 28. Trends Parasitol. 2014. PMID: 24685202 Free PMC article. Review.
-
Omega-3 and omega-6 polyunsaturated fatty acids and their potential therapeutic role in protozoan infections.Front Immunol. 2024 Apr 3;15:1339470. doi: 10.3389/fimmu.2024.1339470. eCollection 2024. Front Immunol. 2024. PMID: 38633251 Free PMC article. Review.
-
Host lipid droplets: An important source of lipids salvaged by the intracellular parasite Toxoplasma gondii.PLoS Pathog. 2017 Jun 1;13(6):e1006362. doi: 10.1371/journal.ppat.1006362. eCollection 2017 Jun. PLoS Pathog. 2017. PMID: 28570716 Free PMC article.
-
PI4-kinase and PfCDPK7 signaling regulate phospholipid biosynthesis in Plasmodium falciparum.EMBO Rep. 2022 Feb 3;23(2):e54022. doi: 10.15252/embr.202154022. Epub 2021 Dec 6. EMBO Rep. 2022. PMID: 34866326 Free PMC article.
References
-
- Hotez PJ, Fenwick A, Savioli L, Molyneux DH. Rescuing the bottom billion through control of neglected tropical diseases. Lancet. 2009;373:1570–5. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
