

Corticosteroids block autophagy protein recruitment in Aspergillus fumigatus phagosomes via targeting dectin-1/Syk kinase signaling
- PMID: 23817424
- PMCID: PMC3883106
- DOI: 10.4049/jimmunol.1300132
Corticosteroids block autophagy protein recruitment in Aspergillus fumigatus phagosomes via targeting dectin-1/Syk kinase signaling
Abstract
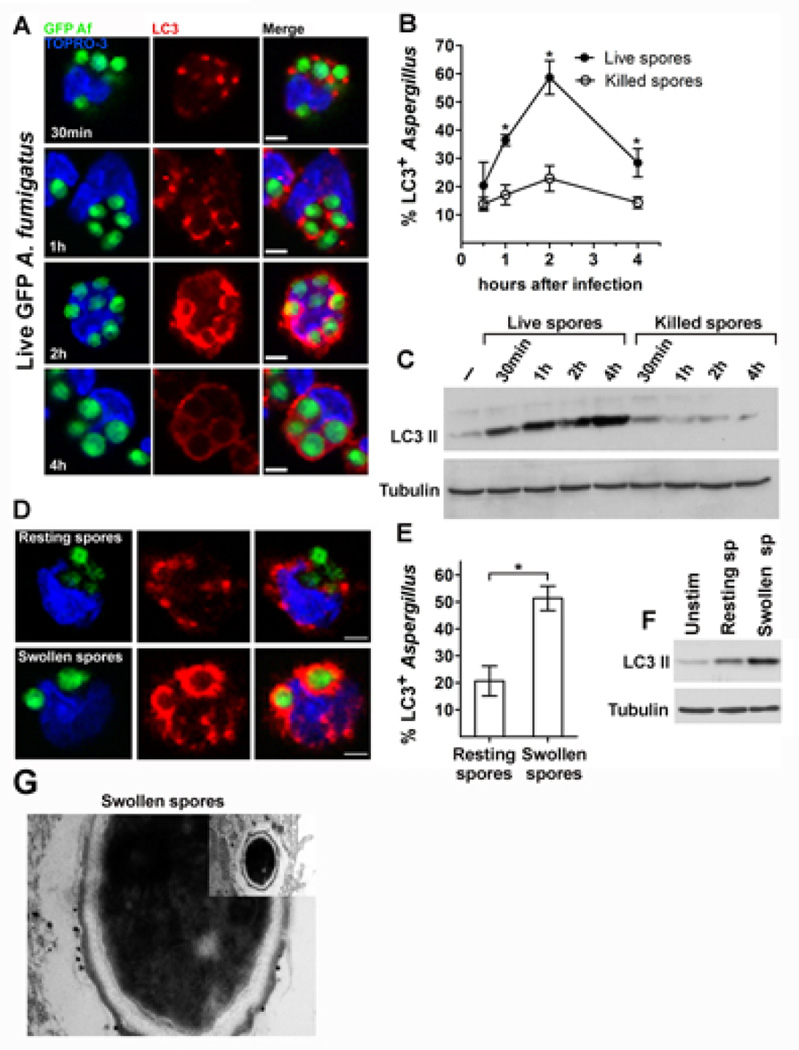
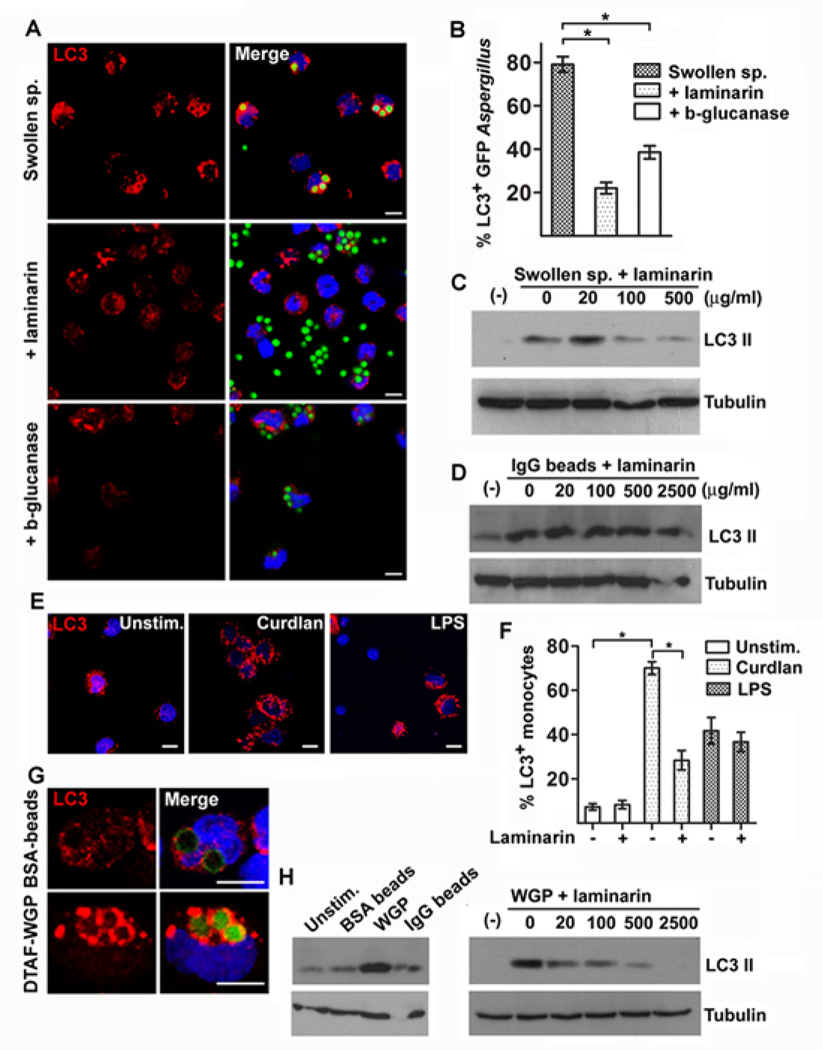
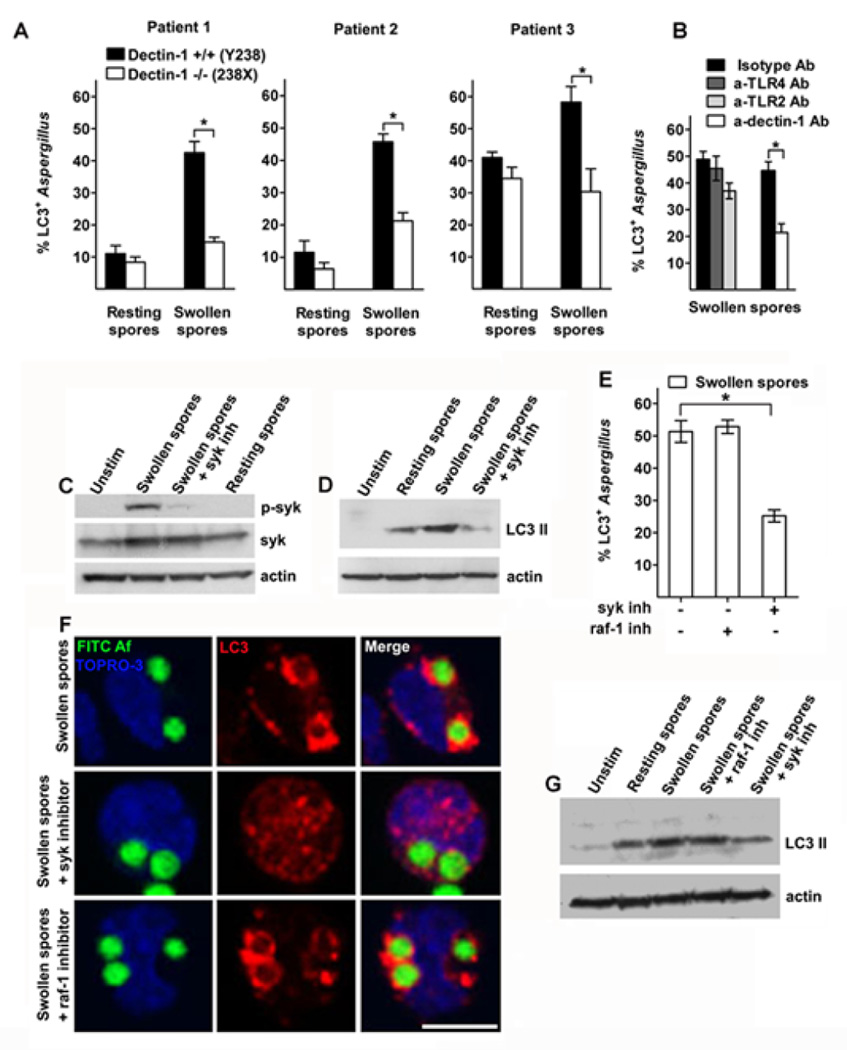
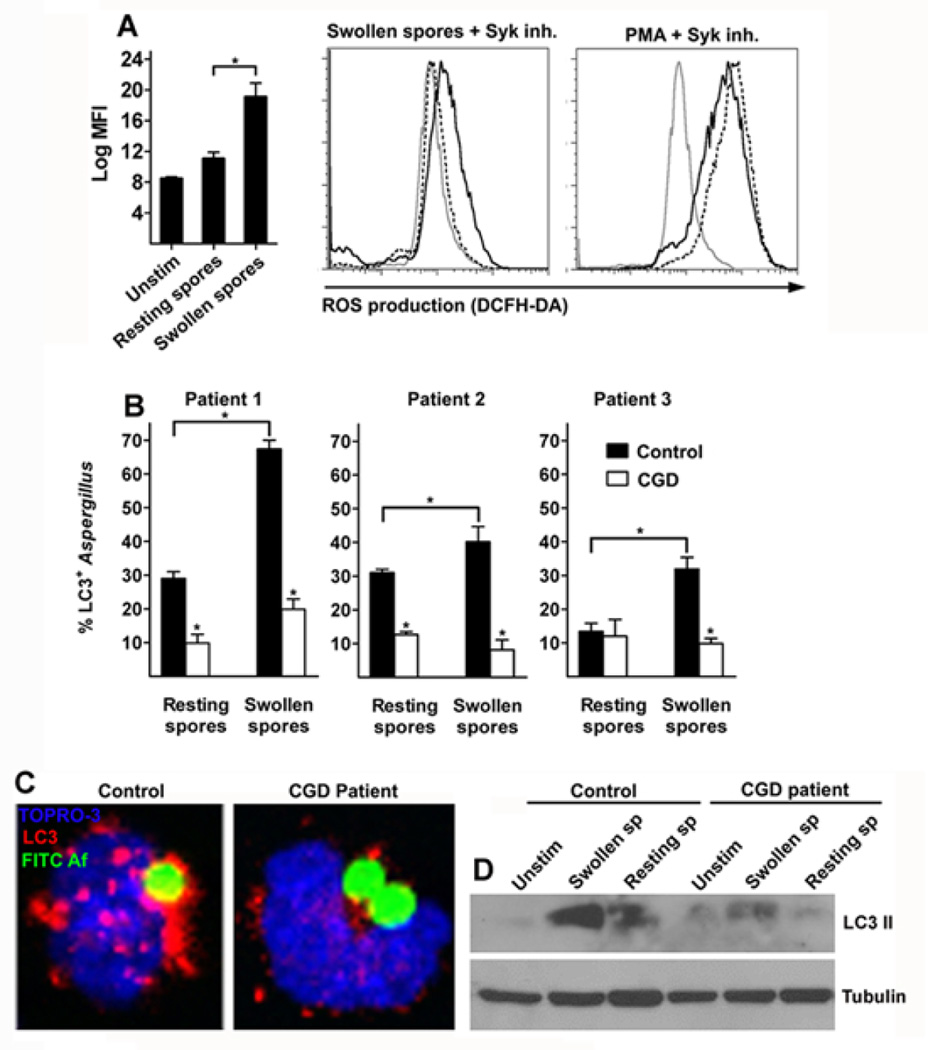
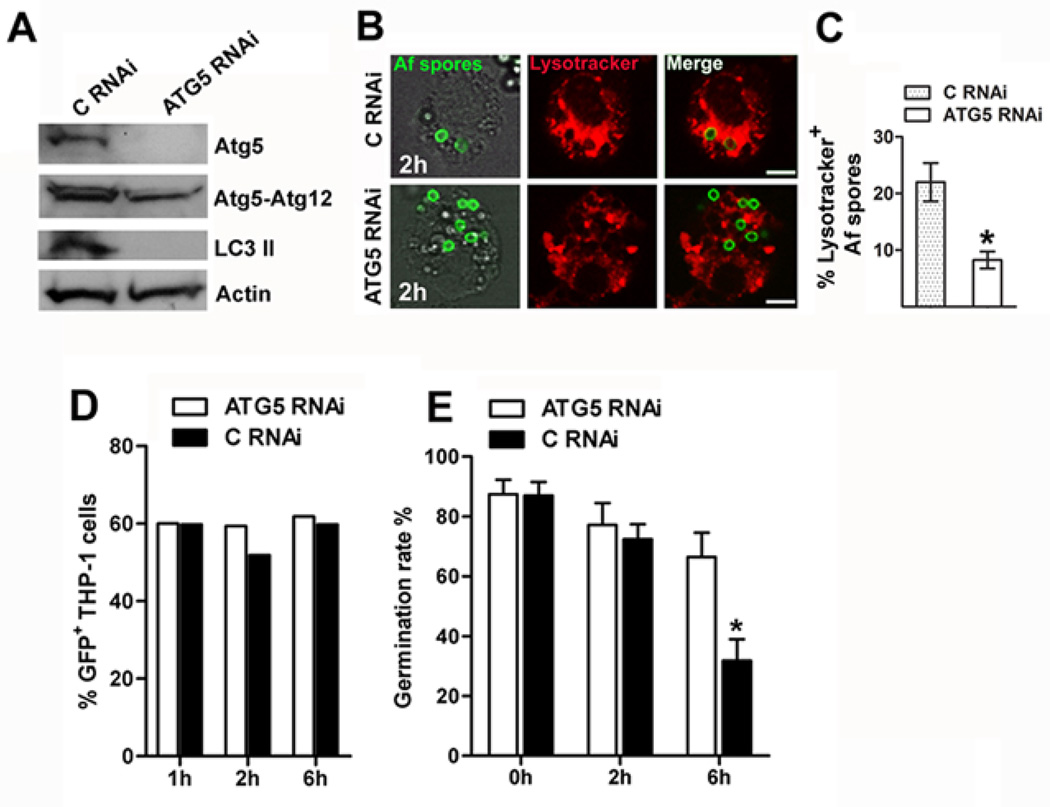
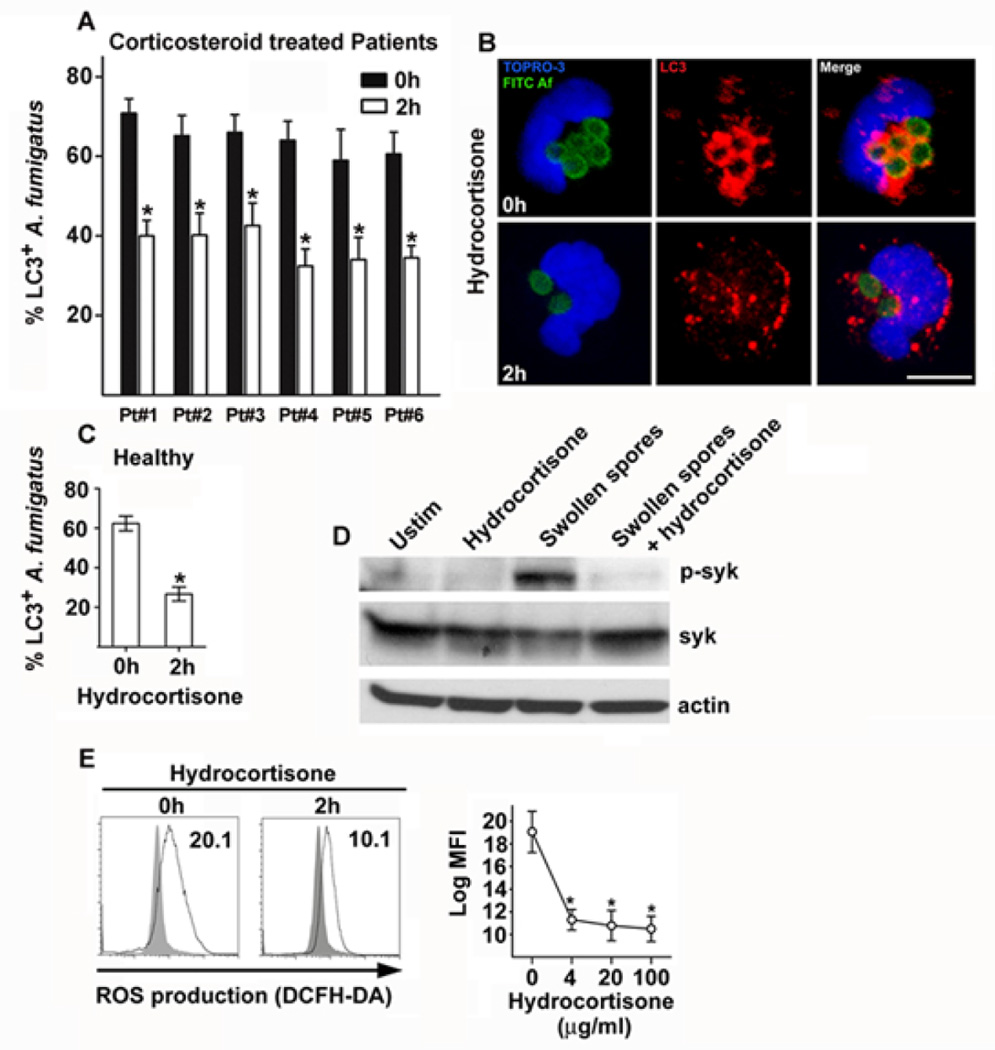
Aspergillus fumigatus is the predominant airborne fungal pathogen in immunocompromised patients. Genetic defects in NADPH oxidase (chronic granulomatous disease [CGD]) and corticosteroid-induced immunosupression lead to impaired killing of A. fumigatus and unique susceptibility to invasive aspergillosis via incompletely characterized mechanisms. Recent studies link TLR activation with phagosome maturation via the engagement of autophagy proteins. In this study, we found that infection of human monocytes with A. fumigatus spores triggered selective recruitment of the autophagy protein LC3 II in phagosomes upon fungal cell wall swelling. This response was induced by surface exposure of immunostimulatory β-glucans and was mediated by activation of the Dectin-1 receptor. LC3 II recruitment in A. fumigatus phagosomes required spleen tyrosine kinase (Syk) kinase-dependent production of reactive oxygen species and was nearly absent in monocytes of patients with CGD. This pathway was important for control of intracellular fungal growth, as silencing of Atg5 resulted in impaired phagosome maturation and killing of A. fumigatus. In vivo and ex vivo administration of corticosteroids blocked LC3 II recruitment in A. fumigatus phagosomes via rapid inhibition of phosphorylation of Src and Syk kinases and downstream production of reactive oxygen species. Our studies link Dectin-1/Syk kinase signaling with autophagy-dependent maturation of A. fumigatus phagosomes and uncover a potential mechanism for development of invasive aspergillosis in the setting of CGD and corticosteroid-induced immunosupression.
Figures
Similar articles
-
Activation of NF-κB and respiratory burst following Aspergillus fumigatus stimulation of macrophages.Immunobiology. 2014 Jan;219(1):25-36. doi: 10.1016/j.imbio.2013.06.013. Epub 2013 Jul 4. Immunobiology. 2014. PMID: 23886693
-
Dectin-1-triggered recruitment of light chain 3 protein to phagosomes facilitates major histocompatibility complex class II presentation of fungal-derived antigens.J Biol Chem. 2012 Oct 5;287(41):34149-56. doi: 10.1074/jbc.M112.382812. Epub 2012 Aug 17. J Biol Chem. 2012. PMID: 22902620 Free PMC article.
-
Dectin-1-dependent LC3 recruitment to phagosomes enhances fungicidal activity in macrophages.J Infect Dis. 2014 Dec 1;210(11):1844-54. doi: 10.1093/infdis/jiu290. Epub 2014 May 19. J Infect Dis. 2014. PMID: 24842831 Free PMC article.
-
LC3-associated phagocytosis: a crucial mechanism for antifungal host defence against Aspergillus fumigatus.Cell Microbiol. 2016 Sep;18(9):1208-16. doi: 10.1111/cmi.12616. Epub 2016 Jul 6. Cell Microbiol. 2016. PMID: 27185357 Review.
-
The role of Dectin-1 in the host defence against fungal infections.Curr Opin Microbiol. 2011 Aug;14(4):392-9. doi: 10.1016/j.mib.2011.07.001. Epub 2011 Jul 29. Curr Opin Microbiol. 2011. PMID: 21803640 Review.
Cited by
-
Insights in Chloroquine Action: Perspectives and Implications in Malaria and COVID-19.Cytometry A. 2020 Sep;97(9):872-881. doi: 10.1002/cyto.a.24190. Epub 2020 Aug 21. Cytometry A. 2020. PMID: 32686260 Free PMC article. Review.
-
Beyond autophagy: LC3-associated phagocytosis and endocytosis.Sci Adv. 2022 Oct 28;8(43):eabn1702. doi: 10.1126/sciadv.abn1702. Epub 2022 Oct 26. Sci Adv. 2022. PMID: 36288309 Free PMC article. Review.
-
Targeting Selective Autophagy as a Therapeutic Strategy for Viral Infectious Diseases.Front Microbiol. 2022 Apr 28;13:889835. doi: 10.3389/fmicb.2022.889835. eCollection 2022. Front Microbiol. 2022. PMID: 35572624 Free PMC article. Review.
-
Targeting autophagy regulation in NLRP3 inflammasome-mediated lung inflammation in COVID-19.Clin Immunol. 2022 Nov;244:109093. doi: 10.1016/j.clim.2022.109093. Epub 2022 Aug 6. Clin Immunol. 2022. PMID: 35944881 Free PMC article. Review.
-
Stimulating the autophagic-lysosomal axis enhances host defense against fungal infection in a zebrafish model of invasive Aspergillosis.Autophagy. 2023 Jan;19(1):324-337. doi: 10.1080/15548627.2022.2090727. Epub 2022 Jul 1. Autophagy. 2023. PMID: 35775203 Free PMC article.
References
-
- Romani L. Immunity to fungal infections. Nat Rev Immunol. 2011;11(4):275–288. - PubMed
-
- Lionakis M, Kontoyiannis DP. Glucocorticoids and invasive fungal infections. Lancet. 2003;362(9398):1828–1838. - PubMed
-
- Cornillet A, Camus C, Nimubona S, et al. Comparison of epidemiological, clinical, and biological features of invasive aspergillosis in neutropenic and nonneutropenic patients: a 6 year survey. Clin Infect Dis. 2006;43(5):577–584. - PubMed
-
- Ader F, Nseir S, Le Berre R, et al. Invasive pulmonary aspergillosis in chronic obstructive pulmonary disease: an emerging fungal pathogen. Clin Microbiol Infect. 2005;11(6):427–429. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous