

Fragile X-associated tremor/ataxia syndrome (FXTAS): pathology and mechanisms
- PMID: 23793382
- PMCID: PMC3904666
- DOI: 10.1007/s00401-013-1138-1
Fragile X-associated tremor/ataxia syndrome (FXTAS): pathology and mechanisms
Abstract
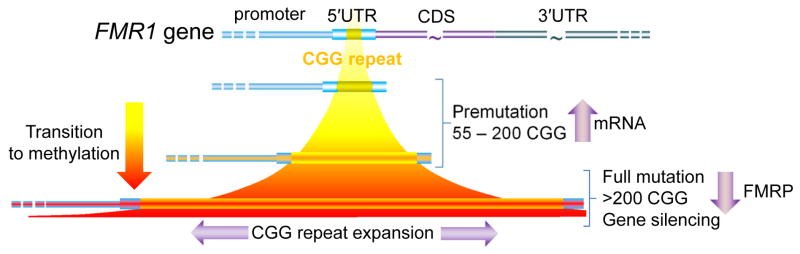
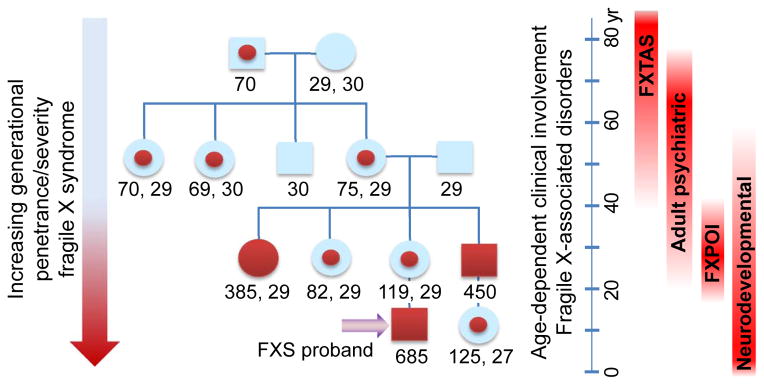
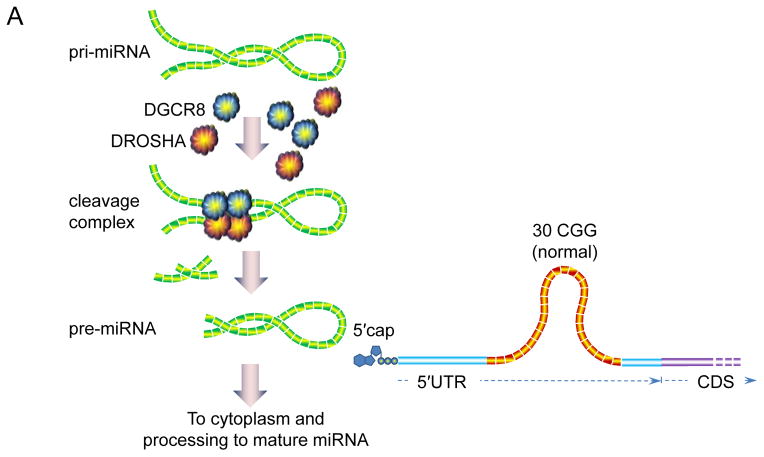
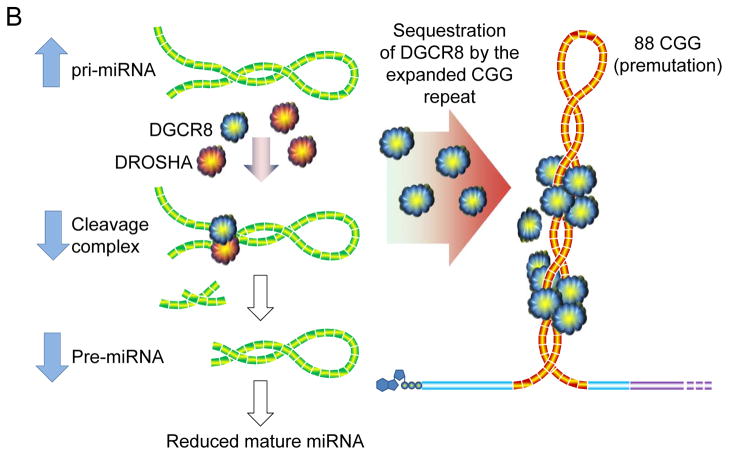
Since its discovery in 2001, our understanding of fragile X-associated tremor/ataxia syndrome (FXTAS) has undergone a remarkable transformation. Initially characterized rather narrowly as an adult-onset movement disorder, the definition of FXTAS is broadening; moreover, the disorder is now recognized as only one facet of a much broader clinical pleiotropy among children and adults who carry premutation alleles of the FMR1 gene. Furthermore, the intranuclear inclusions of FXTAS, once thought to be a CNS-specific marker of the disorder, are now known to be widely distributed in multiple non-CNS tissues; this observation fundamentally changes our concept of the disease, and may provide the basis for understanding the diverse medical problems associated with the premutation. Recent work on the pathogenic mechanisms underlying FXTAS indicates that the origins of the late-onset neurodegenerative disorder actually lie in early development, raising the likelihood that all forms of clinical involvement among premutation carriers have a common underlying mechanistic basis. There has also been great progress in our understanding of the triggering event(s) in FXTAS pathogenesis, which is now thought to involve sequestration of one or more nuclear proteins involved with microRNA biogenesis. Moreover, there is mounting evidence that mitochondrial dysregulation contributes to the decreased cell function and loss of viability, evident in mice even during the neonatal period. Taken together, these recent findings offer hope for early interventions for FXTAS, well before the onset of overt disease, and for the treatment of other forms of clinical involvement among premutation carriers.
Conflict of interest statement
Figures

Similar articles
-
Fragile X-associated tremor/ataxia syndrome: influence of the FMR1 gene on motor fiber tracts in males with normal and premutation alleles.JAMA Neurol. 2013 Aug;70(8):1022-9. doi: 10.1001/jamaneurol.2013.2934. JAMA Neurol. 2013. PMID: 23753897 Free PMC article.
-
Neuropathological, clinical and molecular pathology in female fragile X premutation carriers with and without FXTAS.Genes Brain Behav. 2012 Jul;11(5):577-85. doi: 10.1111/j.1601-183X.2012.00779.x. Epub 2012 Apr 6. Genes Brain Behav. 2012. PMID: 22463693 Free PMC article.
-
What has been learned from mouse models of the Fragile X Premutation and Fragile X-associated tremor/ataxia syndrome?Clin Neuropsychol. 2016 Aug;30(6):960-72. doi: 10.1080/13854046.2016.1158254. Epub 2016 Jun 29. Clin Neuropsychol. 2016. PMID: 27355912 Free PMC article. Review.
-
Volumetric brain changes in females with fragile X-associated tremor/ataxia syndrome (FXTAS).Neurology. 2007 Aug 28;69(9):851-9. doi: 10.1212/01.wnl.0000269781.10417.7b. Neurology. 2007. PMID: 17724287
-
The spectrum of tremor among carriers of the FMR1 premutation with or without the fragile X-associated tremor/ataxia syndrome (FXTAS).Parkinsonism Relat Disord. 2019 Aug;65:32-38. doi: 10.1016/j.parkreldis.2019.05.010. Epub 2019 May 7. Parkinsonism Relat Disord. 2019. PMID: 31126791 Review.
Cited by
-
CGG Repeat-Associated Non-AUG Translation Utilizes a Cap-Dependent Scanning Mechanism of Initiation to Produce Toxic Proteins.Mol Cell. 2016 Apr 21;62(2):314-322. doi: 10.1016/j.molcel.2016.02.034. Epub 2016 Mar 31. Mol Cell. 2016. PMID: 27041225 Free PMC article.
-
Broad autism spectrum and obsessive-compulsive symptoms in adults with the fragile X premutation.Clin Neuropsychol. 2016 Aug;30(6):929-43. doi: 10.1080/13854046.2016.1189536. Epub 2016 Jun 29. Clin Neuropsychol. 2016. PMID: 27355445 Free PMC article.
-
RAN translation at CGG repeats induces ubiquitin proteasome system impairment in models of fragile X-associated tremor ataxia syndrome.Hum Mol Genet. 2015 Aug 1;24(15):4317-26. doi: 10.1093/hmg/ddv165. Epub 2015 May 7. Hum Mol Genet. 2015. PMID: 25954027 Free PMC article.
-
Fragile X spectrum disorders.Intractable Rare Dis Res. 2014 Nov;3(4):134-46. doi: 10.5582/irdr.2014.01022. Intractable Rare Dis Res. 2014. PMID: 25606363 Free PMC article. Review.
-
Composition of the Intranuclear Inclusions of Fragile X-associated Tremor/Ataxia Syndrome.Acta Neuropathol Commun. 2019 Sep 3;7(1):143. doi: 10.1186/s40478-019-0796-1. Acta Neuropathol Commun. 2019. PMID: 31481131 Free PMC article.
References
-
- Adams JS, Adams PE, Nguyen D, Brunberg JA, Tassone F, Zhang W, Koldewyn K, Rivera SM, Grigsby J, Zhang L, DeCarli C, Hagerman PJ, Hagerman RJ. Volumetric brain changes in females with fragile X-associated tremor/ataxia syndrome (FXTAS) Neurology. 2007;69 (9):851–859. doi: 10.1212/01.wnl.0000269781.10417.7b. - DOI - PubMed
-
- Adams PE, Adams JS, Nguyen DV, Hessl D, Brunberg JA, Tassone F, Zhang W, Koldewyn K, Rivera SM, Grigsby J, Zhang L, Decarli C, Hagerman PJ, Hagerman RJ. Psychological symptoms correlate with reduced hippocampal volume in fragile X premutation carriers. Am J Med Genet B Neuropsychiatr Genet. 2010;153B (3):775–785. doi: 10.1002/ajmg.b.31046. - DOI - PMC - PubMed
-
- Apartis E, Blancher A, Meissner WG, Guyant-Marechal L, Maltete D, De Broucker T, Legrand AP, Bouzenada H, Thanh HT, Sallansonnet-Froment M, Wang A, Tison F, Roue-Jagot C, Sedel F, Charles P, Whalen S, Heron D, Thobois S, Poisson A, Lesca G, Ouvrard-Hernandez AM, Fraix V, Palfi S, Habert MO, Gaymard B, Dussaule JC, Pollak P, Vidailhet M, Durr A, Barbot JC, Gourlet V, Brice A, Anheim M. FXTAS: new insights and the need for revised diagnostic criteria. Neurology. 2012;79 (18):1898–1907. doi: 10.1212/WNL.0b013e318271f7ff. - DOI - PubMed
-
- Arocena DG, Iwahashi CK, Won N, Beilina A, Ludwig AL, Tassone F, Schwartz PH, Hagerman PJ. Induction of inclusion formation and disruption of lamin A/C structure by premutation CGG-repeat RNA in human cultured neural cells. Hum Mol Genet. 2005;14 (23):3661–3671. doi: 10.1093/hmg/ddi394. - DOI - PubMed
-
- Ash PE, Bieniek KF, Gendron TF, Caulfield T, Lin WL, Dejesus-Hernandez M, van Blitterswijk MM, Jansen-West K, Paul JW, 3rd, Rademakers R, Boylan KB, Dickson DW, Petrucelli L. Unconventional translation of C9ORF72 GGGGCC expansion generates insoluble polypeptides specific to c9FTD/ALS. Neuron. 2013;77 (4):639–646. doi: 10.1016/j.neuron.2013.02.004. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
