

Substrate binding stabilizes a pre-translocation intermediate in the ATP-binding cassette transport protein MsbA
- PMID: 23766512
- PMCID: PMC3724623
- DOI: 10.1074/jbc.M113.485714
Substrate binding stabilizes a pre-translocation intermediate in the ATP-binding cassette transport protein MsbA
Abstract
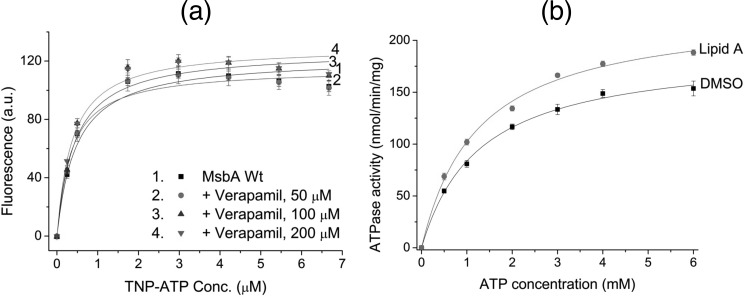
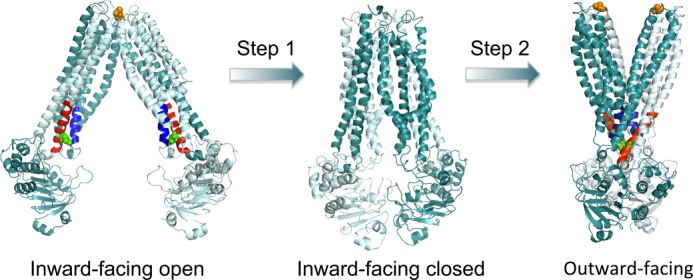
ATP-binding cassette (ABC) transporters belong to one of the largest protein superfamilies that expands from prokaryotes to man. Recent x-ray crystal structures of bacterial and mammalian ABC exporters suggest a common alternating access mechanism of substrate transport, which has also been biochemically substantiated. However, the current model does not yet explain the coupling between substrate binding and ATP hydrolysis that underlies ATP-dependent substrate transport. In our studies on the homodimeric multidrug/lipid A ABC exporter MsbA from Escherichia coli, we performed cysteine cross-linking, fluorescence energy transfer, and cysteine accessibility studies on two reporter positions, near the nucleotide-binding domains and in the membrane domains, for transporter embedded in a biological membrane. Our results suggest for the first time that substrate binding by MsbA stimulates the maximum rate of ATP hydrolysis by facilitating the dimerization of nucleotide-binding domains in a state, which is markedly distinct from the previously described nucleotide-free, inward-facing and nucleotide-bound, outward-facing conformations of ABC exporters and which binds ATP.
Keywords: ABC Transporter; Lipid Transport; Membrane Transport; Multidrug Transporters; Protein Conformation; Substrate-stimulated ATPase.
Figures


Similar articles
-
Dissection of the conformational cycle of the multidrug/lipidA ABC exporter MsbA.Proteins. 2010 Nov 1;78(14):2867-72. doi: 10.1002/prot.22813. Proteins. 2010. PMID: 20715055
-
ATP-dependent substrate transport by the ABC transporter MsbA is proton-coupled.Nat Commun. 2016 Aug 8;7:12387. doi: 10.1038/ncomms12387. Nat Commun. 2016. PMID: 27499013 Free PMC article.
-
Molecular disruption of the power stroke in the ATP-binding cassette transport protein MsbA.J Biol Chem. 2013 Mar 8;288(10):6801-13. doi: 10.1074/jbc.M112.430074. Epub 2013 Jan 10. J Biol Chem. 2013. PMID: 23306205 Free PMC article.
-
Structural basis for the mechanism of ABC transporters.Biochem Soc Trans. 2015 Oct;43(5):889-93. doi: 10.1042/BST20150047. Biochem Soc Trans. 2015. PMID: 26517899 Review.
-
The structures of MsbA: Insight into ABC transporter-mediated multidrug efflux.FEBS Lett. 2006 Feb 13;580(4):1042-8. doi: 10.1016/j.febslet.2005.11.033. Epub 2005 Dec 1. FEBS Lett. 2006. PMID: 16337944 Review.
Cited by
-
Structure of an antibacterial peptide ATP-binding cassette transporter in a novel outward occluded state.Proc Natl Acad Sci U S A. 2014 Jun 24;111(25):9145-50. doi: 10.1073/pnas.1320506111. Epub 2014 Jun 11. Proc Natl Acad Sci U S A. 2014. PMID: 24920594 Free PMC article.
-
Dynamics of ABC Transporter P-glycoprotein in Three Conformational States.Sci Rep. 2019 Oct 22;9(1):15092. doi: 10.1038/s41598-019-50578-2. Sci Rep. 2019. PMID: 31641149 Free PMC article.
-
A Conserved Motif in Intracellular Loop 1 Stabilizes the Outward-Facing Conformation of TmrAB.J Mol Biol. 2021 Aug 6;433(16):166834. doi: 10.1016/j.jmb.2021.166834. Epub 2021 Jan 29. J Mol Biol. 2021. PMID: 33524413 Free PMC article.
-
How to move an amphipathic molecule across a lipid bilayer: different mechanisms for different ABC transporters?Biochem Soc Trans. 2016 Jun 15;44(3):774-82. doi: 10.1042/BST20160040. Biochem Soc Trans. 2016. PMID: 27284041 Free PMC article. Review.
-
Structural Insights into the Lipid A Transport Pathway in MsbA.Structure. 2019 Jul 2;27(7):1114-1123.e3. doi: 10.1016/j.str.2019.04.007. Epub 2019 May 23. Structure. 2019. PMID: 31130486 Free PMC article.
References
-
- Velamakanni S., Yao Y., Gutmann D. A., van Veen H. W. (2008) Multidrug transport by the ABC transporter Sav1866 from Staphylococcus aureus. Biochemistry 47, 9300–9308 - PubMed
-
- Reuter G., Janvilisri T., Venter H., Shahi S., Balakrishnan L., van Veen H. W. (2003) The ATP binding cassette multidrug transporter LmrA and lipid transporter MsbA have overlapping substrate specificities. J. Biol. Chem. 278, 35193–35198 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
