

Splicing functions and global dependency on fission yeast slu7 reveal diversity in spliceosome assembly
- PMID: 23754748
- PMCID: PMC3753915
- DOI: 10.1128/MCB.00007-13
Splicing functions and global dependency on fission yeast slu7 reveal diversity in spliceosome assembly
Abstract
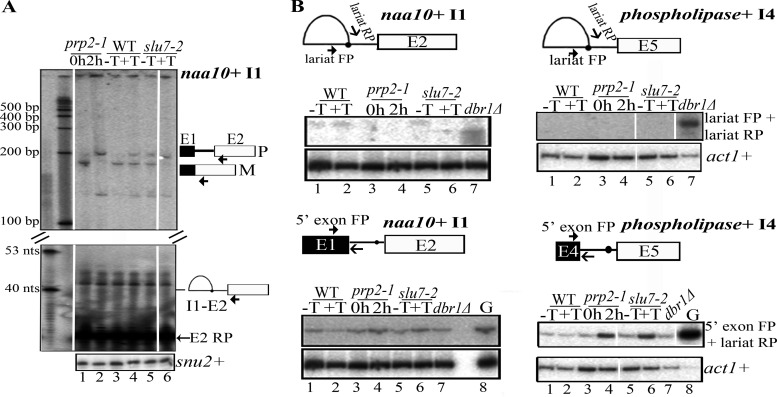
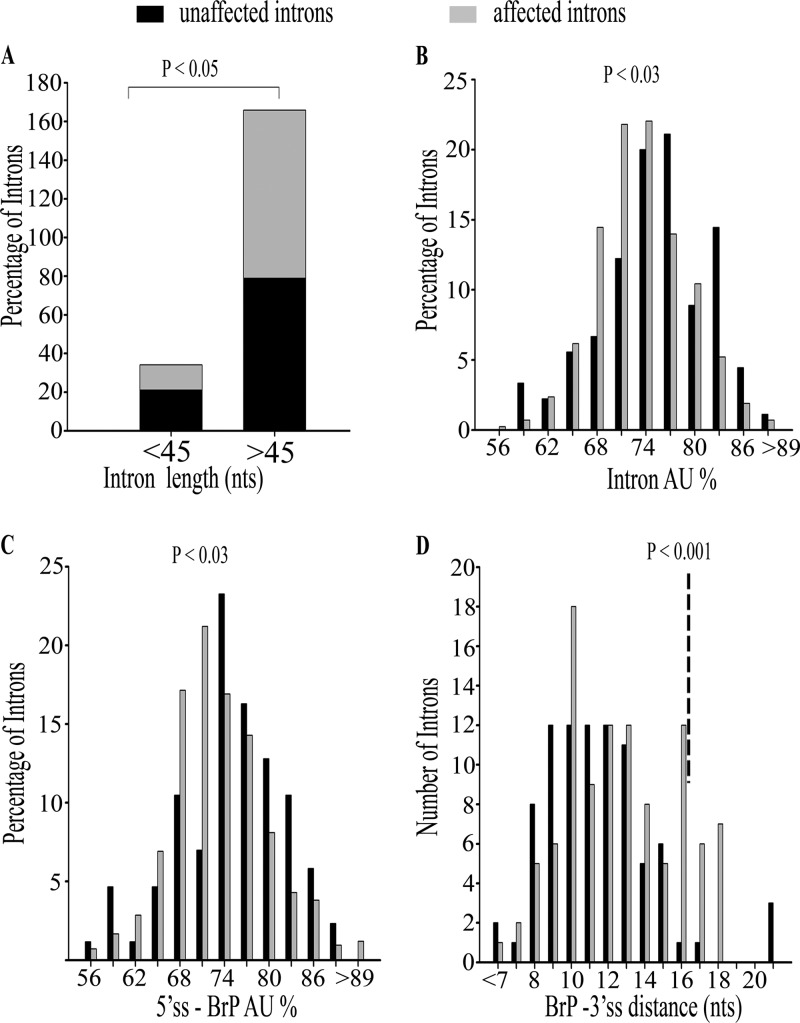
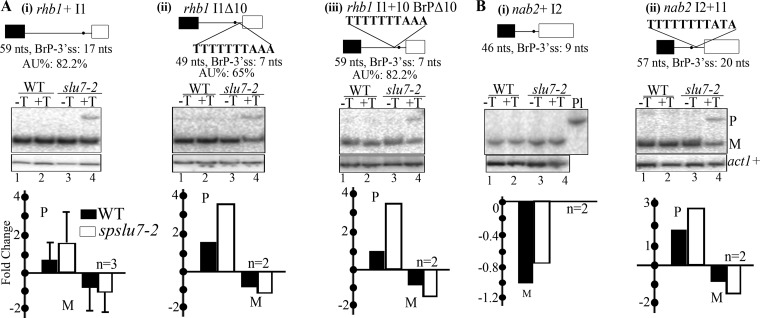
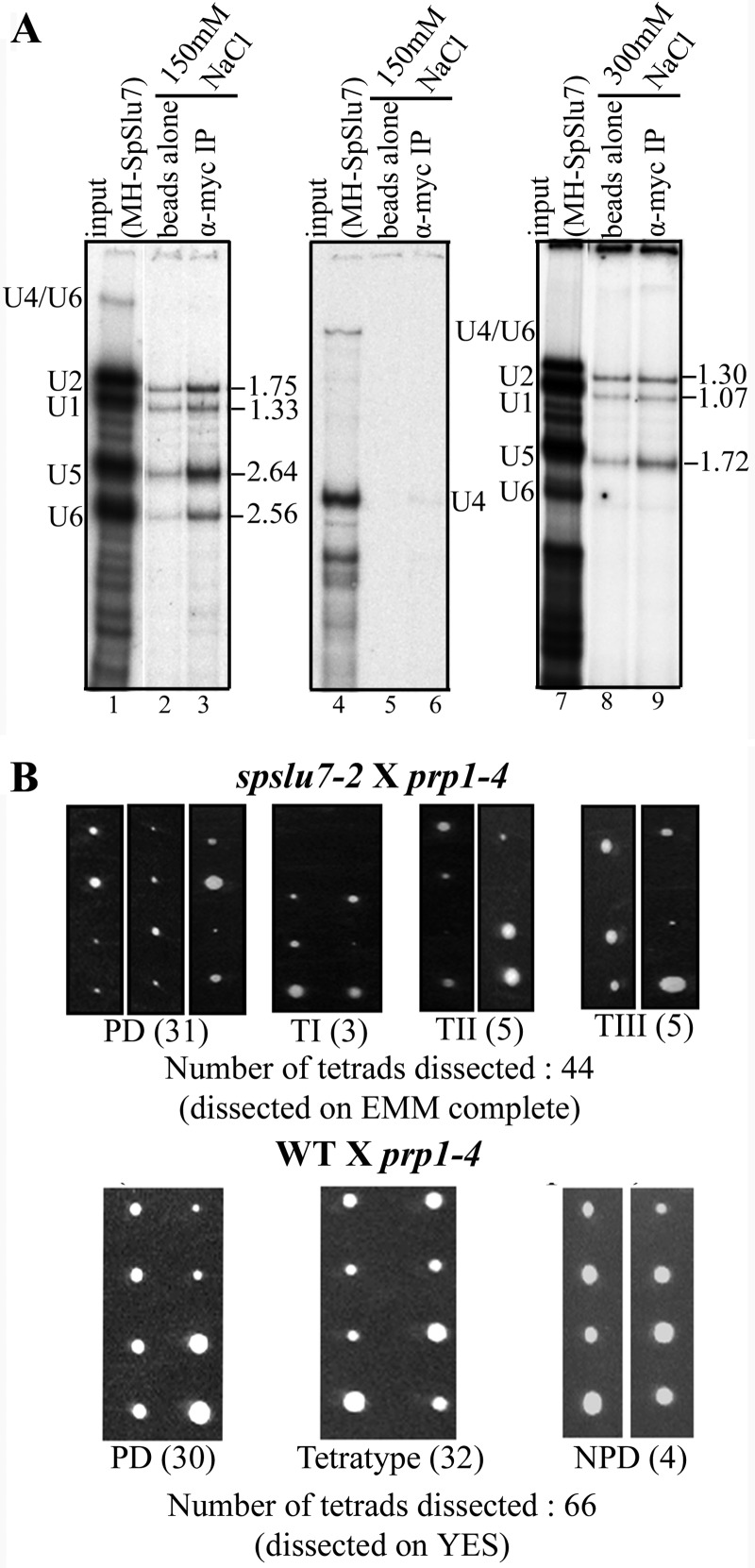
The multiple short introns in Schizosaccharomyces pombe genes with degenerate cis sequences and atypically positioned polypyrimidine tracts make an interesting model to investigate canonical and alternative roles for conserved splicing factors. Here we report functions and interactions of the S. pombe slu7(+) (spslu7(+)) gene product, known from Saccharomyces cerevisiae and human in vitro reactions to assemble into spliceosomes after the first catalytic reaction and to dictate 3' splice site choice during the second reaction. By using a missense mutant of this essential S. pombe factor, we detected a range of global splicing derangements that were validated in assays for the splicing status of diverse candidate introns. We ascribe widespread, intron-specific SpSlu7 functions and have deduced several features, including the branch nucleotide-to-3' splice site distance, intron length, and the impact of its A/U content at the 5' end on the intron's dependence on SpSlu7. The data imply dynamic substrate-splicing factor relationships in multiintron transcripts. Interestingly, the unexpected early splicing arrest in spslu7-2 revealed a role before catalysis. We detected a salt-stable association with U5 snRNP and observed genetic interactions with spprp1(+), a homolog of human U5-102k factor. These observations together point to an altered recruitment and dependence on SpSlu7, suggesting its role in facilitating transitions that promote catalysis, and highlight the diversity in spliceosome assembly.
Figures

Similar articles
-
Genomic mRNA profiling reveals compensatory mechanisms for the requirement of the essential splicing factor U2AF.Mol Cell Biol. 2011 Feb;31(4):652-61. doi: 10.1128/MCB.01000-10. Epub 2010 Dec 13. Mol Cell Biol. 2011. PMID: 21149581 Free PMC article.
-
Role of the ubiquitin-like protein Hub1 in splice-site usage and alternative splicing.Nature. 2011 May 25;474(7350):173-8. doi: 10.1038/nature10143. Nature. 2011. PMID: 21614000 Free PMC article.
-
Genome-wide analysis of pre-mRNA splicing: intron features govern the requirement for the second-step factor, Prp17 in Saccharomyces cerevisiae and Schizosaccharomyces pombe.J Biol Chem. 2004 Dec 10;279(50):52437-46. doi: 10.1074/jbc.M408815200. Epub 2004 Sep 27. J Biol Chem. 2004. PMID: 15452114
-
Multiple genetic and biochemical interactions of Brr2, Prp8, Prp31, Prp1 and Prp4 kinase suggest a function in the control of the activation of spliceosomes in Schizosaccharomyces pombe.Curr Genet. 2005 Sep;48(3):151-61. doi: 10.1007/s00294-005-0013-6. Epub 2005 Oct 12. Curr Genet. 2005. PMID: 16133344 Review.
-
Roles of the U5 snRNP in spliceosome dynamics and catalysis.Biochem Soc Trans. 2004 Dec;32(Pt 6):928-31. doi: 10.1042/BST0320928. Biochem Soc Trans. 2004. PMID: 15506927 Review.
Cited by
-
Cryptococcus neoformans Slu7 ensures nuclear positioning during mitotic progression through RNA splicing.PLoS Genet. 2024 May 20;20(5):e1011272. doi: 10.1371/journal.pgen.1011272. eCollection 2024 May. PLoS Genet. 2024. PMID: 38768219 Free PMC article.
-
Splicing of branchpoint-distant exons is promoted by Cactin, Tls1 and the ubiquitin-fold-activated Sde2.Nucleic Acids Res. 2022 Sep 23;50(17):10000-10014. doi: 10.1093/nar/gkac769. Nucleic Acids Res. 2022. PMID: 36095128 Free PMC article.
-
Early splicing functions of fission yeast Prp16 and its unexpected requirement for gene Silencing is governed by intronic features.RNA Biol. 2019 Jun;16(6):754-769. doi: 10.1080/15476286.2019.1585737. Epub 2019 Mar 20. RNA Biol. 2019. PMID: 30810475 Free PMC article.
-
Prp4 Kinase Grants the License to Splice: Control of Weak Splice Sites during Spliceosome Activation.PLoS Genet. 2016 Jan 5;12(1):e1005768. doi: 10.1371/journal.pgen.1005768. eCollection 2016 Jan. PLoS Genet. 2016. PMID: 26730850 Free PMC article.
-
Functions for fission yeast splicing factors SpSlu7 and SpPrp18 in alternative splice-site choice and stress-specific regulated splicing.PLoS One. 2017 Dec 13;12(12):e0188159. doi: 10.1371/journal.pone.0188159. eCollection 2017. PLoS One. 2017. PMID: 29236736 Free PMC article.
References
-
- Burge CB, Tuschl TH, Sharp PA. 1999. Splicing of precursors to mRNAs by the spliceosome, p 525–560 In Gesteland RF, Cech TR, Atkins JF. (ed), RNA World II. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases