

The LXR-IDOL axis defines a clathrin-, caveolae-, and dynamin-independent endocytic route for LDLR internalization and lysosomal degradation
- PMID: 23733886
- PMCID: PMC3708367
- DOI: 10.1194/jlr.M037713
The LXR-IDOL axis defines a clathrin-, caveolae-, and dynamin-independent endocytic route for LDLR internalization and lysosomal degradation
Abstract
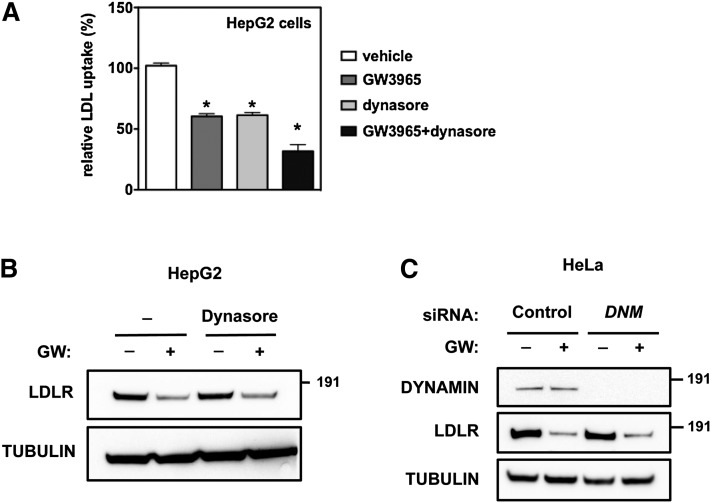
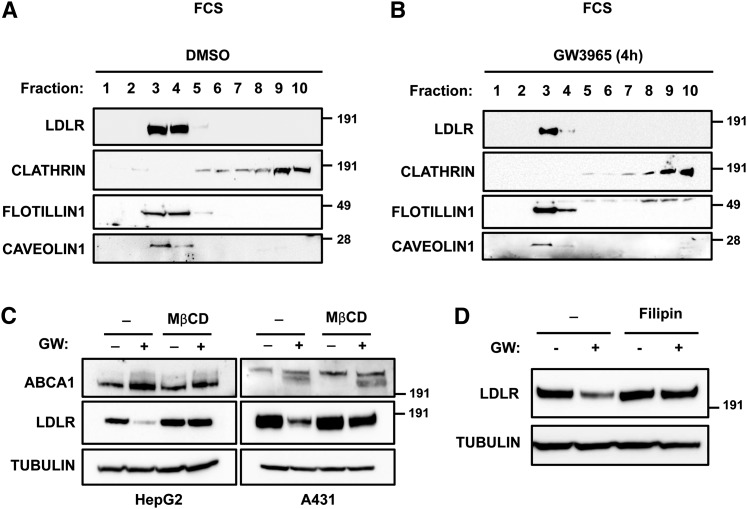
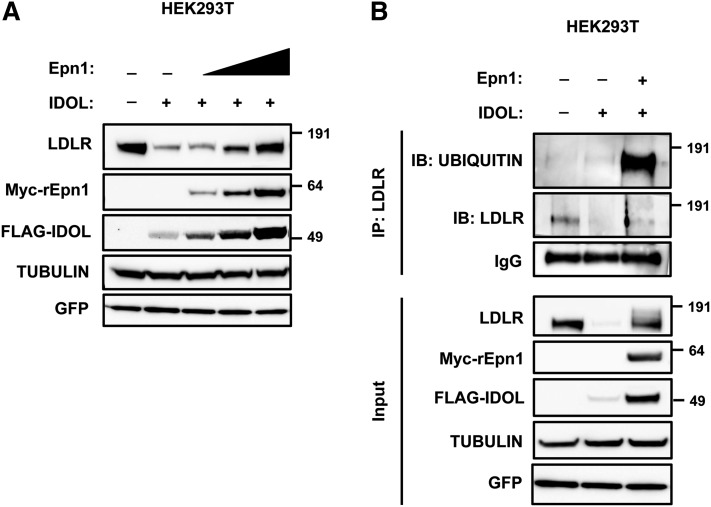
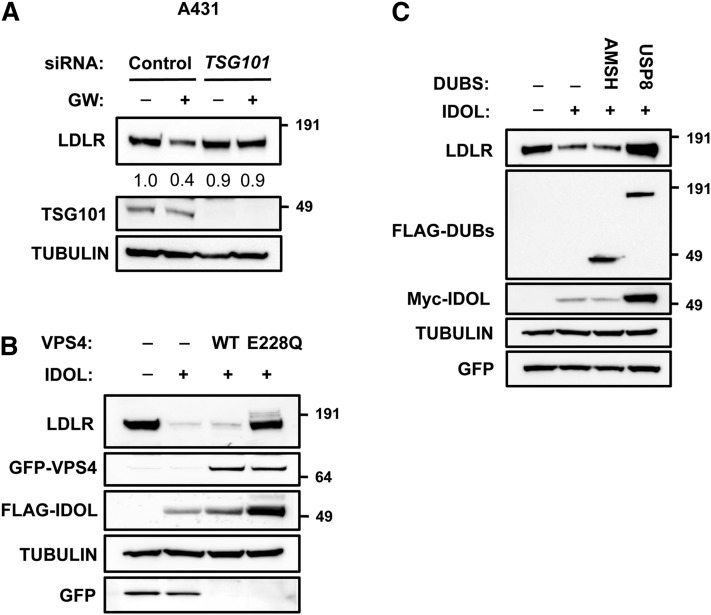
Low density lipoprotein (LDL) cholesterol is taken up into cells via clathrin-mediated endocytosis of the LDL receptor (LDLR). Following dissociation of the LDLR-LDL complex, LDL is directed to lysosomes whereas the LDLR recycles to the plasma membrane. Activation of the sterol-sensing nuclear receptors liver X receptors (LXRs) enhances degradation of the LDLR. This depends on the LXR target gene inducible degrader of the LDLR (IDOL), an E3-ubiquitin ligase that promotes ubiquitylation and lysosomal degradation of the LDLR. How ubiquitylation of the LDLR by IDOL controls its endocytic trafficking is currently unknown. Using genetic- and pharmacological-based approaches coupled to functional assessment of LDL uptake, we show that the LXR-IDOL axis targets a LDLR pool present in lipid rafts. IDOL-dependent internalization of the LDLR is independent of clathrin, caveolin, macroautophagy, and dynamin. Rather, it depends on the endocytic protein epsin. Consistent with LDLR ubiquitylation acting as a sorting signal, degradation of the receptor can be blocked by perturbing the endosomal sorting complex required for transport (ESCRT) or by USP8, a deubiquitylase implicated in sorting ubiquitylated cargo to multivesicular bodies. In summary, we provide evidence for the existence of an LXR-IDOL-mediated internalization pathway for the LDLR that is distinct from that used for lipoprotein uptake.
Keywords: E3-ubiquitin ligase; endocytosis; epsins; inducible degrader of low density lipoprotein receptor; lipoprotein receptors; liver X receptor; low density lipoprotein receptor.
Figures
Similar articles
-
IDOL stimulates clathrin-independent endocytosis and multivesicular body-mediated lysosomal degradation of the low-density lipoprotein receptor.Mol Cell Biol. 2013 Apr;33(8):1503-14. doi: 10.1128/MCB.01716-12. Epub 2013 Feb 4. Mol Cell Biol. 2013. PMID: 23382078 Free PMC article.
-
Deubiquitylase Inhibition Reveals Liver X Receptor-independent Transcriptional Regulation of the E3 Ubiquitin Ligase IDOL and Lipoprotein Uptake.J Biol Chem. 2016 Feb 26;291(9):4813-25. doi: 10.1074/jbc.M115.698688. Epub 2015 Dec 30. J Biol Chem. 2016. PMID: 26719329 Free PMC article.
-
Distinct functional domains contribute to degradation of the low density lipoprotein receptor (LDLR) by the E3 ubiquitin ligase inducible Degrader of the LDLR (IDOL).J Biol Chem. 2011 Aug 26;286(34):30190-9. doi: 10.1074/jbc.M111.249557. Epub 2011 Jul 6. J Biol Chem. 2011. PMID: 21734303 Free PMC article.
-
Feedback regulation of cholesterol uptake by the LXR-IDOL-LDLR axis.Arterioscler Thromb Vasc Biol. 2012 Nov;32(11):2541-6. doi: 10.1161/ATVBAHA.112.250571. Epub 2012 Aug 30. Arterioscler Thromb Vasc Biol. 2012. PMID: 22936343 Free PMC article. Review.
-
Post-transcriptional regulation of lipoprotein receptors by the E3-ubiquitin ligase inducible degrader of the low-density lipoprotein receptor.Curr Opin Lipidol. 2012 Jun;23(3):213-219. doi: 10.1097/MOL.0b013e3283532947. Curr Opin Lipidol. 2012. PMID: 22510808 Review.
Cited by
-
The hepatic WASH complex is required for efficient plasma LDL and HDL cholesterol clearance.JCI Insight. 2019 Jun 6;4(11):e126462. doi: 10.1172/jci.insight.126462. eCollection 2019 Jun 6. JCI Insight. 2019. PMID: 31167970 Free PMC article.
-
β-Estradiol results in a proprotein convertase subtilisin/kexin type 9-dependent increase in low-density lipoprotein receptor levels in human hepatic HuH7 cells.FEBS J. 2015 Jul;282(14):2682-96. doi: 10.1111/febs.13309. Epub 2015 May 18. FEBS J. 2015. PMID: 25913303 Free PMC article.
-
Myeloid-Specific Deletion of Epsins 1 and 2 Reduces Atherosclerosis by Preventing LRP-1 Downregulation.Circ Res. 2019 Feb 15;124(4):e6-e19. doi: 10.1161/CIRCRESAHA.118.313028. Circ Res. 2019. PMID: 30595089 Free PMC article.
-
Liver X receptors in lipid metabolism: opportunities for drug discovery.Nat Rev Drug Discov. 2014 Jun;13(6):433-44. doi: 10.1038/nrd4280. Epub 2014 May 16. Nat Rev Drug Discov. 2014. PMID: 24833295 Review.
-
Hepatitis C Virus Uses Host Lipids to Its Own Advantage.Metabolites. 2021 Apr 27;11(5):273. doi: 10.3390/metabo11050273. Metabolites. 2021. PMID: 33925362 Free PMC article. Review.
References
-
- Brown M. S., Goldstein J. L. 1986. A receptor-mediated pathway for cholesterol homeostasis. Science. 232: 34–47 - PubMed
-
- Garcia C. K. 2001. Autosomal recessive hypercholesterolemia caused by mutations in a putative LDL receptor adaptor protein. Science. 292: 1394–1398 - PubMed
-
- Chen W. J., Goldstein J. L., Brown M. S. 1990. NPXY, a sequence often found in cytoplasmic tails, is required for coated pit-mediated internalization of the low density lipoprotein receptor. J. Biol. Chem. 265: 3116–3123 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
