

The human PDZome: a gateway to PSD95-Disc large-zonula occludens (PDZ)-mediated functions
- PMID: 23722234
- PMCID: PMC3769332
- DOI: 10.1074/mcp.O112.021022
The human PDZome: a gateway to PSD95-Disc large-zonula occludens (PDZ)-mediated functions
Abstract
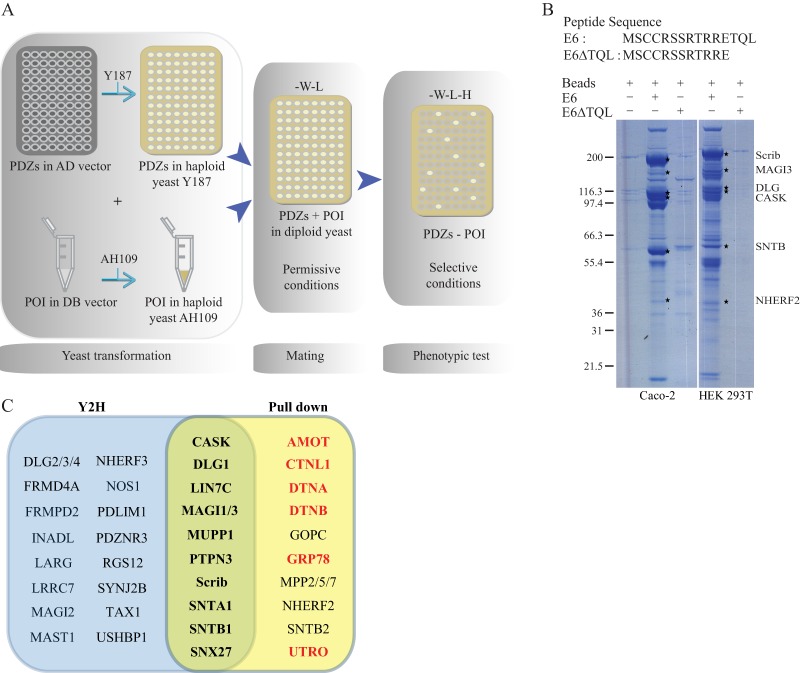
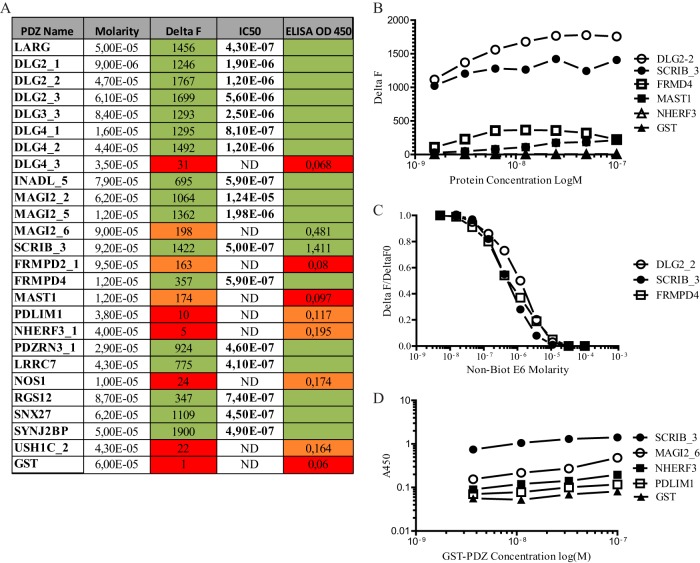
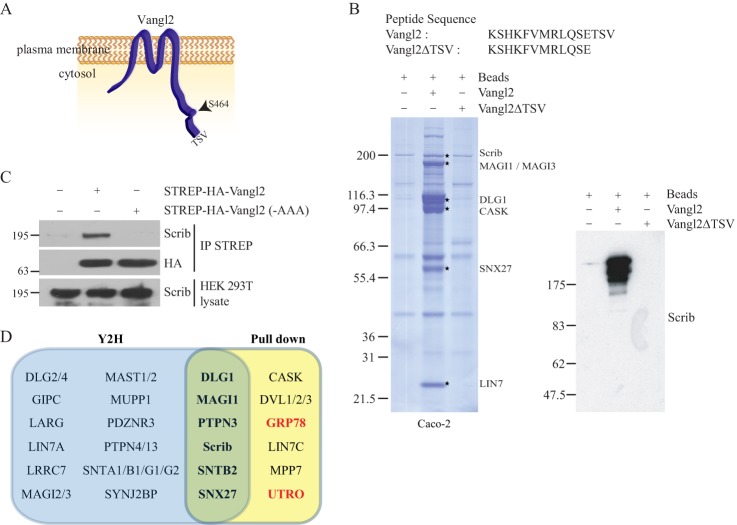
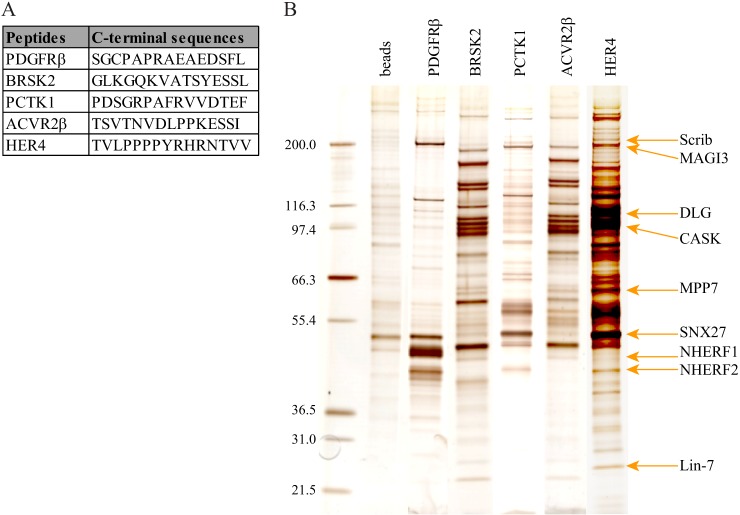
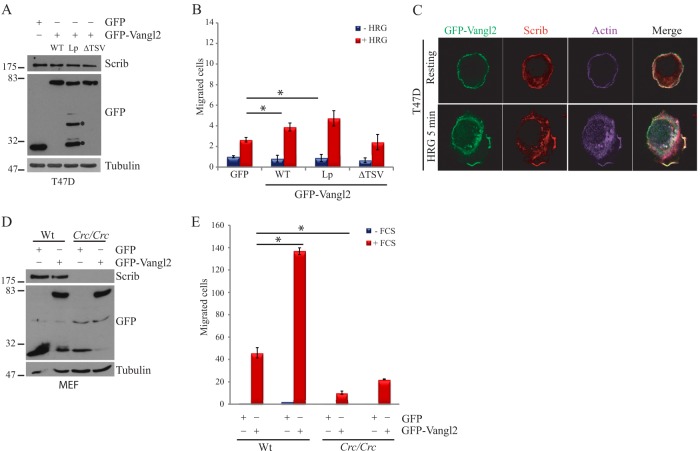
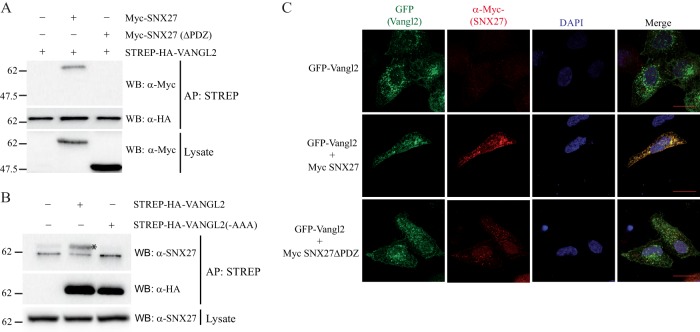
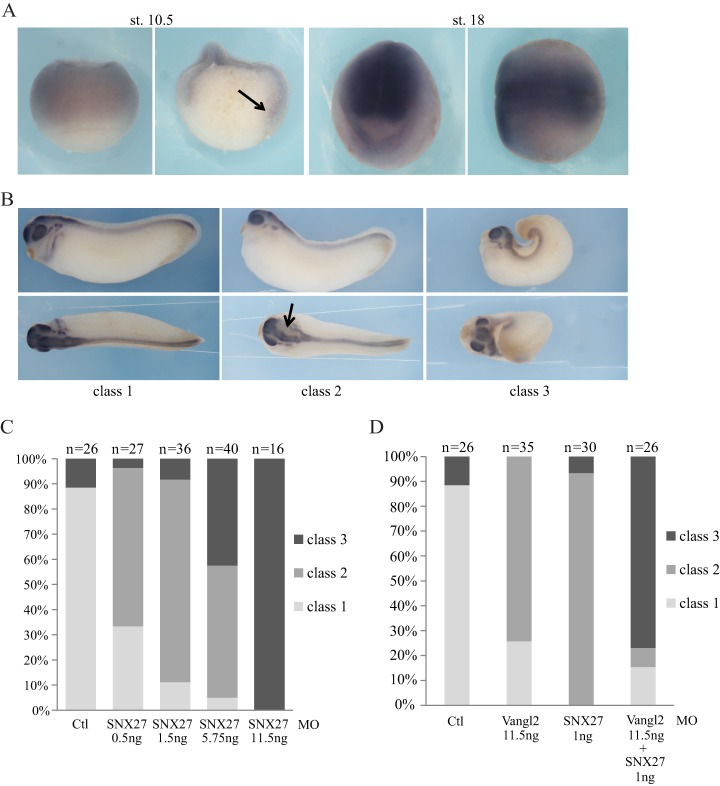
Protein-protein interactions organize the localization, clustering, signal transduction, and degradation of cellular proteins and are therefore implicated in numerous biological functions. These interactions are mediated by specialized domains able to bind to modified or unmodified peptides present in binding partners. Among the most broadly distributed protein interaction domains, PSD95-disc large-zonula occludens (PDZ) domains are usually able to bind carboxy-terminal sequences of their partners. In an effort to accelerate the discovery of PDZ domain interactions, we have constructed an array displaying 96% of the human PDZ domains that is amenable to rapid two-hybrid screens in yeast. We have demonstrated that this array can efficiently identify interactions using carboxy-terminal sequences of PDZ domain binders such as the E6 oncoviral protein and protein kinases (PDGFRβ, BRSK2, PCTK1, ACVR2B, and HER4); this has been validated via mass spectrometry analysis. Taking advantage of this array, we show that PDZ domains of Scrib and SNX27 bind to the carboxy-terminal region of the planar cell polarity receptor Vangl2. We also have demonstrated the requirement of Scrib for the promigratory function of Vangl2 and described the morphogenetic function of SNX27 in the early Xenopus embryo. The resource presented here is thus adapted for the screen of PDZ interactors and, furthermore, should facilitate the understanding of PDZ-mediated functions.
Figures
Similar articles
-
The Human PDZome 2.0: Characterization of a New Resource to Test for PDZ Interactions by Yeast Two-Hybrid.Membranes (Basel). 2023 Aug 17;13(8):737. doi: 10.3390/membranes13080737. Membranes (Basel). 2023. PMID: 37623798 Free PMC article.
-
Structural basis of the human Scribble-Vangl2 association in health and disease.Biochem J. 2021 Apr 16;478(7):1321-1332. doi: 10.1042/BCJ20200816. Biochem J. 2021. PMID: 33684218 Free PMC article.
-
Identification of PDZ Interactions by Proteomic Peptide Phage Display.Methods Mol Biol. 2021;2256:41-60. doi: 10.1007/978-1-0716-1166-1_3. Methods Mol Biol. 2021. PMID: 34014515
-
Mechanism and role of PDZ domains in signaling complex assembly.J Cell Sci. 2001 Sep;114(Pt 18):3219-31. doi: 10.1242/jcs.114.18.3219. J Cell Sci. 2001. PMID: 11591811 Review.
-
Peptide Targeting of PDZ-Dependent Interactions as Pharmacological Intervention in Immune-Related Diseases.Molecules. 2021 Oct 21;26(21):6367. doi: 10.3390/molecules26216367. Molecules. 2021. PMID: 34770776 Free PMC article. Review.
Cited by
-
The Known and Potential Intersections of Rab-GTPases in Human Papillomavirus Infections.Front Cell Dev Biol. 2019 Aug 14;7:139. doi: 10.3389/fcell.2019.00139. eCollection 2019. Front Cell Dev Biol. 2019. PMID: 31475144 Free PMC article. Review.
-
RNA interference screening identifies a novel role for PCTK1/CDK16 in medulloblastoma with c-Myc amplification.Oncotarget. 2015 Jan 1;6(1):116-29. doi: 10.18632/oncotarget.2699. Oncotarget. 2015. PMID: 25402633 Free PMC article.
-
Viral PDZ Binding Motifs Influence Cell Behavior Through the Interaction with Cellular Proteins Containing PDZ Domains.Methods Mol Biol. 2021;2256:217-236. doi: 10.1007/978-1-0716-1166-1_13. Methods Mol Biol. 2021. PMID: 34014525 Review.
-
Vangl as a Master Scaffold for Wnt/Planar Cell Polarity Signaling in Development and Disease.Front Cell Dev Biol. 2022 May 11;10:887100. doi: 10.3389/fcell.2022.887100. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35646914 Free PMC article. Review.
-
PRICKLE1 promotes gastric cancer metastasis by activating mTOR signaling.Am J Transl Res. 2021 May 15;13(5):4266-4280. eCollection 2021. Am J Transl Res. 2021. PMID: 34150013 Free PMC article.
References
-
- Pawson T., Gish G. D., Nash P. (2001) SH2 domains, interaction modules and cellular wiring. Trends Cell Biol. 11, 504–511 - PubMed
-
- Pawson T. (2007) Dynamic control of signaling by modular adaptor proteins. Curr. Opin. Cell Biol. 19, 112–116 - PubMed
-
- Nourry C., Grant S. G., Borg J. P. (2003) PDZ domain proteins: plug and play! Sci. STKE 2003, RE7. - PubMed
-
- Tonikian R., Zhang Y., Sazinsky S. L., Currell B., Yeh J. H., Reva B., Held H. A., Appleton B. A., Evangelista M., Wu Y., Xin X., Chan A. C., Seshagiri S., Lasky L. A., Sander C., Boone C., Bader G. D., Sidhu S. S. (2008) A specificity map for the PDZ domain family. PLoS Biol. 6, e239. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
