

Review
doi: 10.1101/gad.215137.113.
Transcriptional elongation checkpoint control in development and disease
Affiliations
- PMID: 23699407
- PMCID: PMC3672643
- DOI: 10.1101/gad.215137.113
Item in Clipboard
Review
Transcriptional elongation checkpoint control in development and disease
Genes Dev.
.
Abstract
Transcriptional elongation control by RNA polymerase II and its associated factors has taken center stage as a process essential for the regulation of gene expression throughout development. In this review, we analyze recent findings on the identification of factors functioning in the regulation of the transcriptional elongation checkpoint control (TECC) stage of gene expression and how the factors' misregulation is associated with disease pathogenesis, including cancer.
Keywords: RNA polymerase II; cancer; chromatin; elongation; transcription.
Figures
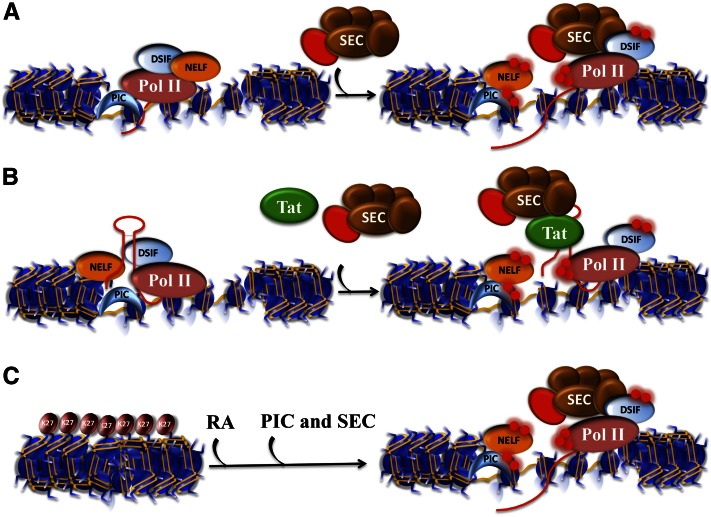
Transcriptional activation with and without paused Pol II. (A) The classic example of transcriptional regulation by paused Pol II was demonstrated for the Hsp70 gene in Drosophila. Pol II is initially recruited to the gene promoter by the PIC (represented here with a TBP-shaped crescent). (Left panel) DSIF cooperates with NELF to keep Pol II within the first 40–70 nt of the transcription start site (TSS). (Right panel) Upon heat shock, the P-TEFb kinase (red sphere), as a component of the SEC, phosphorylates both DSIF and the Pol II CTD to allow the release from NELF, which allows productive elongation by Pol II, while DSIF becomes a positive factor for processivity. (B) Paused Pol II and its release by SEC have been extensively studied during transactivation of the HIV-1 provirus by the HIV-1 encoded Tat protein. Early in the viral life cycle, provirus transcription is inefficient until enough Tat protein has accumulated. Tat binds to a stem–loop structure early in the first 80 nt of the transcript from which it recruits SEC to cause the release of Pol II into productive transcription. (C) Transcriptional induction without paused Pol II. Recent studies in mouse ES cells revealed that one of the most rapidly induced genes in response to differentiation signals, Cyp26a1, lacks paused Pol II in the ES cell state and is covered by H3K27me3. In the presence of retinoic acid (RA), the PIC, Pol II can quickly initiate transcription, and in the presence of SEC, rapid transcriptional induction is ensued (see also Fig. 4A,B).
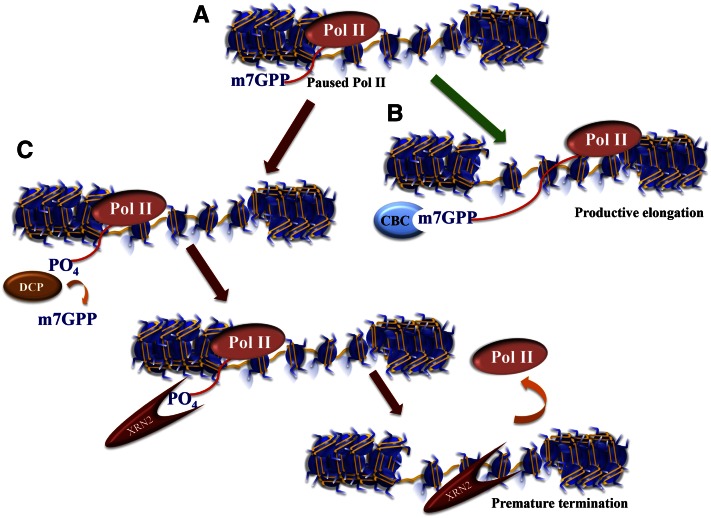
Multiple ways to regulate paused Pol II. Recent studies have revealed other mechanisms to regulate paused Pol II in addition to the DSIF/NELF pause–release mechanism shown in Figure 1. (A) Paused Pol II is a nonproductive state producing short ∼50-nt transcripts that have already been modified with a methyl-7-guanosine cap (m7GPP) protecting the 5′ end of the mRNA from exonucleolytic cleavage. (B) Normally, the cap-binding protein complex (CBC) recognizes the m7G cap and stays with the transcript to facilitate RNA processing and the export of the RNA to the cytoplasm. (C) A decapping enzyme (DCP) can remove the m7G cap, thereby restoring the 5′ phosphate of the RNA, which allows the RNA to be degraded by the XRN2 exonuclease. The 5′ -to-3′ processivity of the XRN2 exonuclease helps push the stalling Pol II off the transcript, resulting in transcription termination. Note that an entry point for XRN2-dependent Pol II termination can also come about from any endonucleolytic cleavage event, such as when the miRNA processing factor Drosha cleaves the stem–loop structure of the TAR region of the HIV-1 transcript (also see Fig. 1).
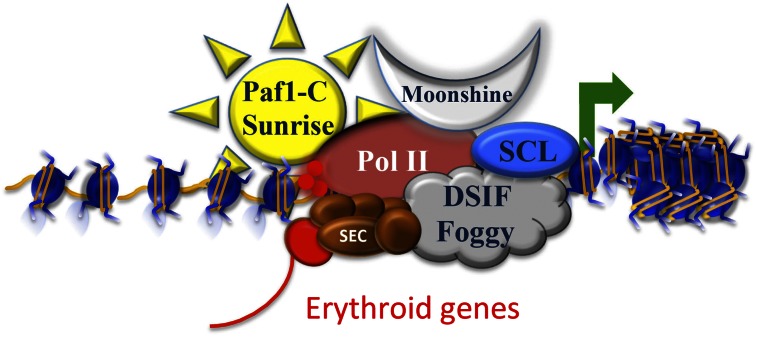
Regulating paused Pol II in the context of chromatin. TIF1γ associates with histone chaperones and elongation factors to stimulate gene expression during erythropoiesis. Mutations in moonshine, encoding zebrafish TIF1γ, could be suppressed by mutations in sunrise, encoding the PAF-c subunit CDC73, and by mutations in foggy, which is an allele of the DSIF subunit SPT5, which loses its ability to maintain the paused Pol II state. The SCL complex, containing the basic helix–loop–helix (bHLH) SCL transcription factor, could recruit TIF1γ and the histone chaperones SPT6 and SPT16/FACT to remodel nucleosomes to facilitate Pol II release. The most active form of P-TEFb (red sphere) within SEC is involved in Pol II CTD phosphorylation and the release from the paused state. These findings implicate transcription elongation as a major regulatory step for the control of erythroid gene expression.
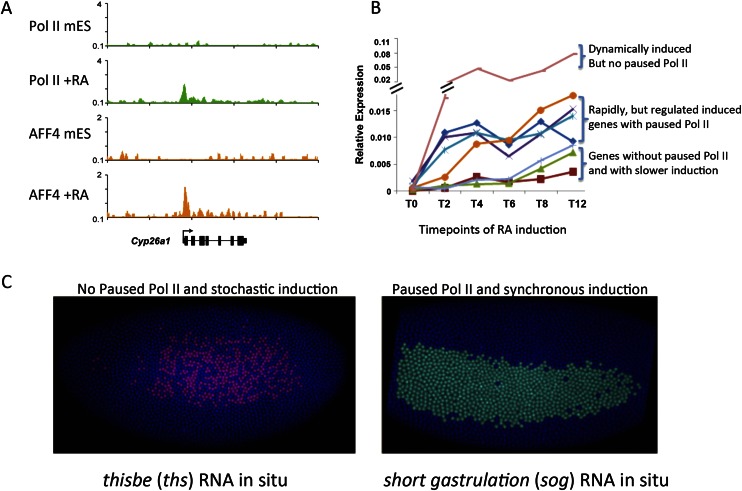
Paused Pol II in equitable and synchronous transcriptional induction. (A) Rapid induction of gene expression can occur without prior paused Pol II. In mouse ES cells (mES), Pol II is absent from the promoter of Cyp26a1. Upon retinoic acid (RA) treatment, Pol II is recruited to Cyp26a1 along with the SEC component AFF4 (Lin et al. 2011). (B) Time course of transcriptional induction in retinoic acid-treated mouse ES cells. Cyp26a1 (pink line/top bracket), which A shows lacks paused Pol II in the ES state, is the most rapidly (dynamically) induced gene in response to retinoic acid treatment. Many genes that are rapidly induced by retinoic acid treatment had paused Pol II in mouse ES cells, although they are induced in a more regulated or equitable manner (middle bracket). This study demonstrated that paused Pol II is not a prerequisite for rapid transcriptional induction (Lin et al. 2011). (C) Paused Pol II correlates with the synchronous induction of genes in early Drosophila embryogenesis. Two genes, thisbe (ths) and short gastrulation (sog), can be distinguished by the absence of paused Pol II in very early embryos. (Left panel) ths in situ hybridization shows that ths, which lacked paused Pol II prior to induction, has a stochastic induction with a nonuniform pattern of expression. (Right panel) A gene with paused Pol II in the early embryo is sog, which, like ths, is induced in the neurogenic ectoderm but has a more uniform induction pattern. Therefore, Boettiger and Levine (2009) propose that the prevalence of paused Pol II on developmentally regulated genes is to ensure a well-regulated, less stochastic expression during embryogenesis. A and B are modified with permission from Genes & Development (Lin et al. 2011). C is modified from Levine 2011 with permission from Elsevier (© 2011).
Similar articles
-
The super elongation complex (SEC) family in transcriptional control.Nat Rev Mol Cell Biol. 2012 Sep;13(9):543-7. doi: 10.1038/nrm3417. Epub 2012 Aug 16. Nat Rev Mol Cell Biol. 2012. PMID: 22895430 Review.
-
Negative elongation factor (NELF) coordinates RNA polymerase II pausing, premature termination, and chromatin remodeling to regulate HIV transcription.J Biol Chem. 2013 Sep 6;288(36):25995-26003. doi: 10.1074/jbc.M113.496489. Epub 2013 Jul 24. J Biol Chem. 2013. PMID: 23884411 Free PMC article.
-
Sub1 associates with Spt5 and influences RNA polymerase II transcription elongation rate.Mol Biol Cell. 2012 Nov;23(21):4297-312. doi: 10.1091/mbc.E12-04-0331. Epub 2012 Sep 12. Mol Biol Cell. 2012. PMID: 22973055 Free PMC article.
-
An mRNA Capping Enzyme Targets FACT to the Active Gene To Enhance the Engagement of RNA Polymerase II into Transcriptional Elongation.Mol Cell Biol. 2017 Jun 15;37(13):e00029-17. doi: 10.1128/MCB.00029-17. Print 2017 Jul 1. Mol Cell Biol. 2017. PMID: 28396559 Free PMC article.
-
A role for histone acetylation in regulating transcription elongation.Transcription. 2018;9(4):225-232. doi: 10.1080/21541264.2017.1394423. Epub 2017 Dec 8. Transcription. 2018. PMID: 29219750 Free PMC article. Review.
Cited by
-
Stably paused genes revealed through inhibition of transcription initiation by the TFIIH inhibitor triptolide.Genes Dev. 2015 Jan 1;29(1):39-47. doi: 10.1101/gad.246173.114. Genes Dev. 2015. PMID: 25561494 Free PMC article.
-
Epigenetics of hematopoiesis and hematological malignancies.Genes Dev. 2016 Sep 15;30(18):2021-2041. doi: 10.1101/gad.284109.116. Genes Dev. 2016. PMID: 27798847 Free PMC article. Review.
-
Global unleashing of transcription elongation waves in response to genotoxic stress restricts somatic mutation rate.Nat Commun. 2017 Dec 12;8(1):2076. doi: 10.1038/s41467-017-02145-4. Nat Commun. 2017. PMID: 29233992 Free PMC article.
-
The plant RNA polymerase II elongation complex: A hub coordinating transcript elongation and mRNA processing.Transcription. 2018;9(2):117-122. doi: 10.1080/21541264.2017.1356902. Epub 2017 Oct 4. Transcription. 2018. PMID: 28886274 Free PMC article.
-
PAF1, a Molecular Regulator of Promoter-Proximal Pausing by RNA Polymerase II.Cell. 2015 Aug 27;162(5):1003-15. doi: 10.1016/j.cell.2015.07.042. Epub 2015 Aug 13. Cell. 2015. PMID: 26279188 Free PMC article.
References
-
- Aso T, Lane WS, Conaway JW, Conaway RC 1995. Elongin (SIII): A multisubunit regulator of elongation by RNA polymerase II. Science 269: 1439–1443 - PubMed
-
- Bartholomeeusen K, Xiang Y, Fujinaga K, Peterlin BM 2012. Bromodomain and extra-terminal (BET) bromodomain inhibition activate transcription via transient release of positive transcription elongation factor b (P-TEFb) from 7SK small nuclear ribonucleoprotein. J Biol Chem 287: 36609–36616 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources