

Review
doi: 10.1038/nrm3591.
Molecular regulation of stem cell quiescence
Affiliations
- PMID: 23698583
- PMCID: PMC3808888
- DOI: 10.1038/nrm3591
Item in Clipboard
Review
Molecular regulation of stem cell quiescence
Nat Rev Mol Cell Biol.
2013 Jun.
Abstract
Subsets of mammalian adult stem cells reside in the quiescent state for prolonged periods of time. This state, which is reversible, has long been viewed as dormant and with minimal basal activity. Recent advances in adult stem cell isolation have provided insights into the epigenetic, transcriptional and post-transcriptional control of quiescence and suggest that quiescence is an actively maintained state in which signalling pathways are involved in maintaining a poised state that allows rapid activation. Deciphering the molecular mechanisms regulating adult stem cell quiescence will increase our understanding of tissue regeneration mechanisms and how they are dysregulated in pathological conditions and in ageing.
Figures
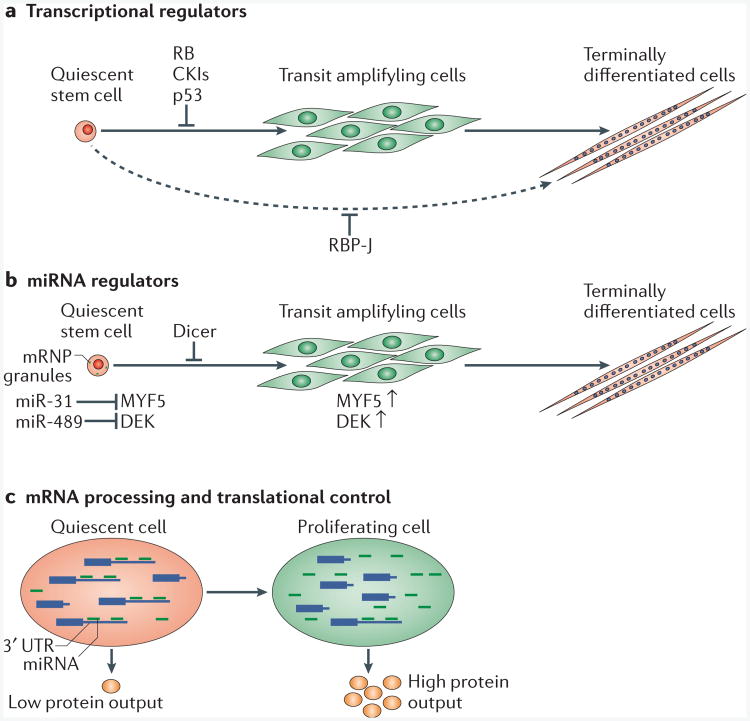
Recent data suggest that the state of quiescence is actively regulated by different molecular mechanisms. a | Quiescence regulators such as RB, cyclin-dependent kinase inhibitors (CKIs) and p53 negatively regulate the activation of quiescent stem cells. In quiescent muscle stem cells, the loss of the downstream effector of the Notch signalling pathway, RBP-J, promotes spontaneous activation and terminal differentiation, in some cases without cell division. b | Quiescent stem cells are actively regulated by post-transcriptional mechanisms. The loss of the microRNA (miRNA)-processing enzyme Dicer or specific miRNAs promotes quiescent stem cell activation. c | Differential mRNA processing alters the susceptibility of mRNAs to miRNA regulation. In quiescent cells, distal polyadenylation signals (PASs) are used to generate mRNA transcripts with long 3′ untranslated regions (3′ UTRs). In proliferating cells, proximal PASs are used, which decrease the number of miRNA target sites on the transcripts, allowing some transcripts to escape miRNA-mediated inhibition and leading to increased protein expression.
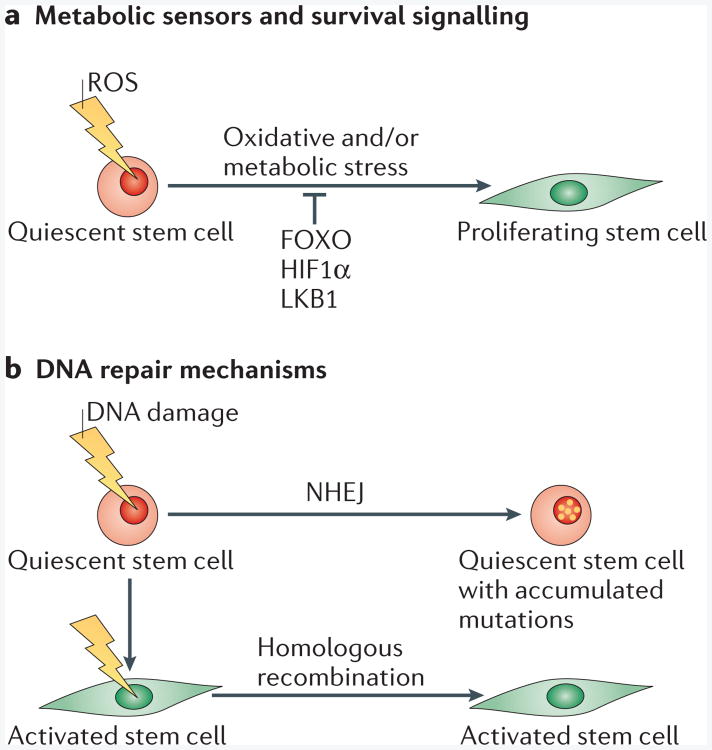
a| Metabolic sensors an d effectors such as forkhead box O (FOXO), hypoxia-inducible factor 1α (HIF1α) and liver kinase B1 (LKB1) are expressed in quiescent stem cells. Expression of these molecules is essential for metabolic functions and survival of quiescent stem cells in adverse environments. These factors protect quiescent stem cells from oxidative stress caused by the accumulation of reactive oxygen species (ROS). Quiescent stem cells devoid of these pathways have an increased propensity to become activated and fail to maintain the stem cell pool. b | Non-homologous end-joining (NHEJ) and homologous recombination are pathways that repair DNA double-strand breaks (DSBs). Homologous recombination is a high-fidelity mechanism that uses homologous templates as guides for DSB repair, whereas NHEJ directly ligates the ends of the DSBs. DSBs that have been repaired by NHEJ can be imprecise when the overhangs at the DSBs are not compatible. In quiescent stem cells, the error-prone NHEJ mechanism is used for DSB repair, which suggests that DNA mutations accumulate in these cells throughout their life. MYF5, myogenic factor 5.
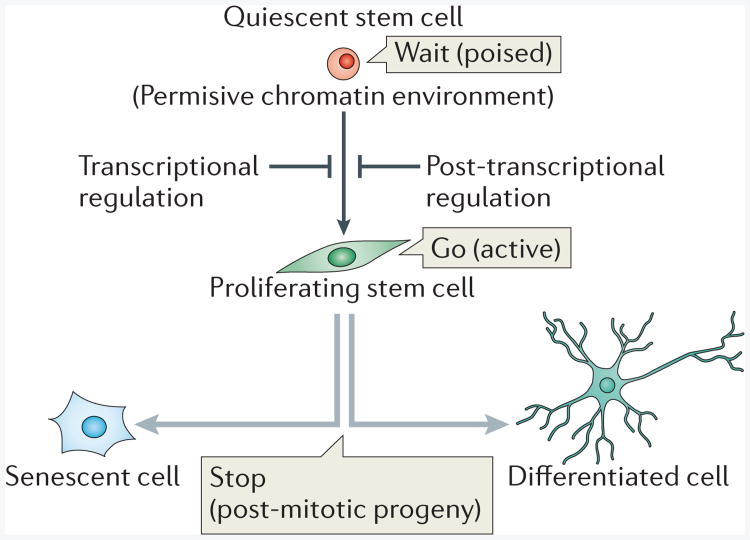
A proposed model of how the quiescent state of a stem cell constitutes a poised state for activation. Quiescent stem cells are actively regulated at the epigenetic, transcriptional and post-transcriptional level. The epigenetic landscape keeps the chromatin in a permissive state, which allows rapid transcriptional activation. Additional layers of transcriptional and post-transcriptional control safeguard quiescent stem cells to enable precise stem cell activation when necessary.
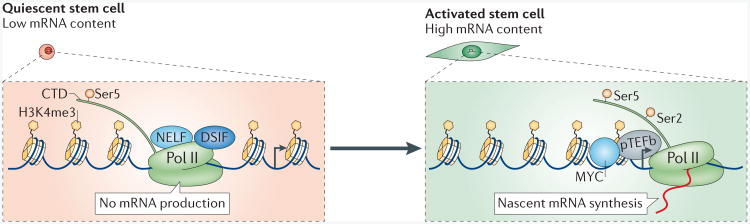
Current data suggest that RNA polymerase II (Pol II) is paused at transcription start sites where histone H3 is trimethylated at Lys 4 (H3K4me3; which is a permissive histone mark). Quiescent stem cells have low mRNA content. The carboxy-terminal domain (CTD) of PolII is phosphorylated at Ser5 but not the Ser2, which indicates transcriptional initiation but not transcriptional elongation. Many types of quiescent stem cells lack Ser2 phosphorylation at the Pol II CTD, which suggests that transcriptional elongation does not occur in these cells, and hence the mRNA levels are low. In quiescent stem cells, Pol II is associated with the negative elongation factors DSIF (DRB sensitivity inducing factor) and NELF. Upon stem cell activation, phosphorylation of DSIF, of NELF (negative elongation factor) and of Ser2 of the Pol II CTD, in combination with the expression of MYC, which is recruited to promoters together with transcription elongation factors (such as pTEFb (positive transcription elongation factor b)), transform the state of promoter proximal pausing into productive elongation and lead to mRNA synthesis.
Similar articles
-
Mechanisms, Hallmarks, and Implications of Stem Cell Quiescence.Stem Cell Reports. 2019 Jun 11;12(6):1190-1200. doi: 10.1016/j.stemcr.2019.05.012. Stem Cell Reports. 2019. PMID: 31189093 Free PMC article. Review.
-
Molecular Regulation of Cellular Quiescence: A Perspective from Adult Stem Cells and Its Niches.Methods Mol Biol. 2018;1686:1-25. doi: 10.1007/978-1-4939-7371-2_1. Methods Mol Biol. 2018. PMID: 29030809 Review.
-
Stem Cell Quiescence: Dynamism, Restraint, and Cellular Idling.Cell Stem Cell. 2019 Feb 7;24(2):213-225. doi: 10.1016/j.stem.2019.01.001. Cell Stem Cell. 2019. PMID: 30735649 Free PMC article. Review.
-
Cellular mechanisms of somatic stem cell aging.Curr Top Dev Biol. 2014;107:405-38. doi: 10.1016/B978-0-12-416022-4.00014-7. Curr Top Dev Biol. 2014. PMID: 24439814 Free PMC article. Review.
-
Fine-tuned Rest: Unveiling the Regulatory Landscape of Adult Quiescent Neural Stem Cells.Neuroscience. 2023 Aug 10;525:26-37. doi: 10.1016/j.neuroscience.2023.07.003. Epub 2023 Jul 10. Neuroscience. 2023. PMID: 37437796 Review.
Cited by
-
The transcription factor Prox1 is essential for satellite cell differentiation and muscle fibre-type regulation.Nat Commun. 2016 Oct 12;7:13124. doi: 10.1038/ncomms13124. Nat Commun. 2016. PMID: 27731315 Free PMC article.
-
Autophagy is a key factor in maintaining the regenerative capacity of muscle stem cells by promoting quiescence and preventing senescence.Autophagy. 2016;12(4):617-8. doi: 10.1080/15548627.2016.1158373. Autophagy. 2016. PMID: 27050452 Free PMC article.
-
Mitochondria as Signaling Organelles Control Mammalian Stem Cell Fate.Cell Stem Cell. 2021 Mar 4;28(3):394-408. doi: 10.1016/j.stem.2021.02.011. Cell Stem Cell. 2021. PMID: 33667360 Free PMC article. Review.
-
p53 Inhibits Bmi-1-driven Self-Renewal and Defines Salivary Gland Cancer Stemness.Clin Cancer Res. 2022 Nov 1;28(21):4757-4770. doi: 10.1158/1078-0432.CCR-22-1357. Clin Cancer Res. 2022. PMID: 36048559 Free PMC article.
-
Odyssey of human dental pulp stem cells and their remarkable ability to survive in extremely adverse conditions.Front Physiol. 2015 Mar 26;6:99. doi: 10.3389/fphys.2015.00099. eCollection 2015. Front Physiol. 2015. PMID: 25859225 Free PMC article. No abstract available.
References
-
- Weissman IL. Stem cells: units of development, units of regeneration, and units in evolution. Cell. 2000;100:157–168. - PubMed
-
- Orford KW, Scadden DT. Deconstructing stem cell self-renewal: genetic insights into cell-cycle regulation. Nature Rev Genet. 2008;9:115–128. - PubMed
-
- Howard A, Pelc SR. Synthesis of deoxyribonucleic acid in normal and irradiated cells and its relation to chromosome breakage. Hered (Lond) [Suppl] 1953;6:261–273.
-
- Baserga R. Biochemistry of the cell cycle: a review. Cell Prolifer. 1968;1:167–191.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
