

T cell antigen receptor activation and actin cytoskeleton remodeling
- PMID: 23680625
- PMCID: PMC3877165
- DOI: 10.1016/j.bbamem.2013.05.004
T cell antigen receptor activation and actin cytoskeleton remodeling
Abstract
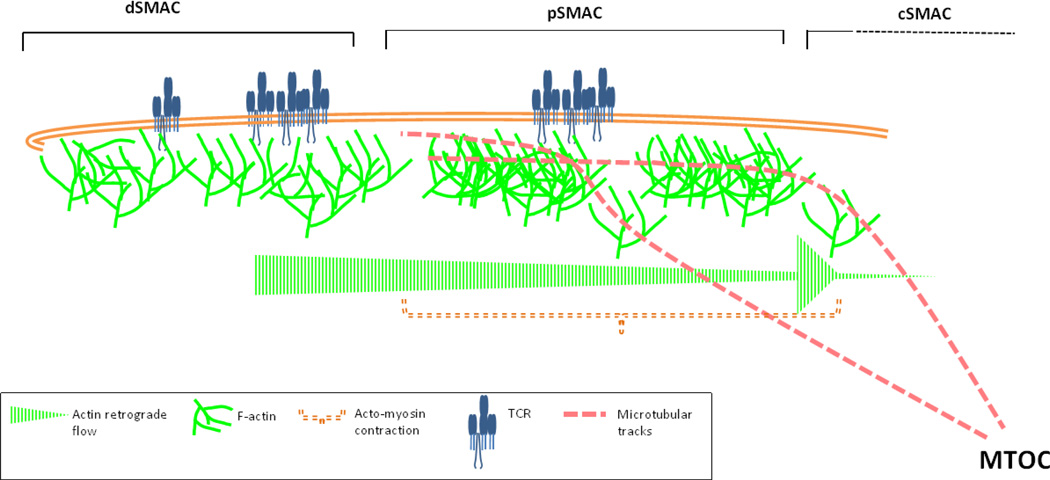
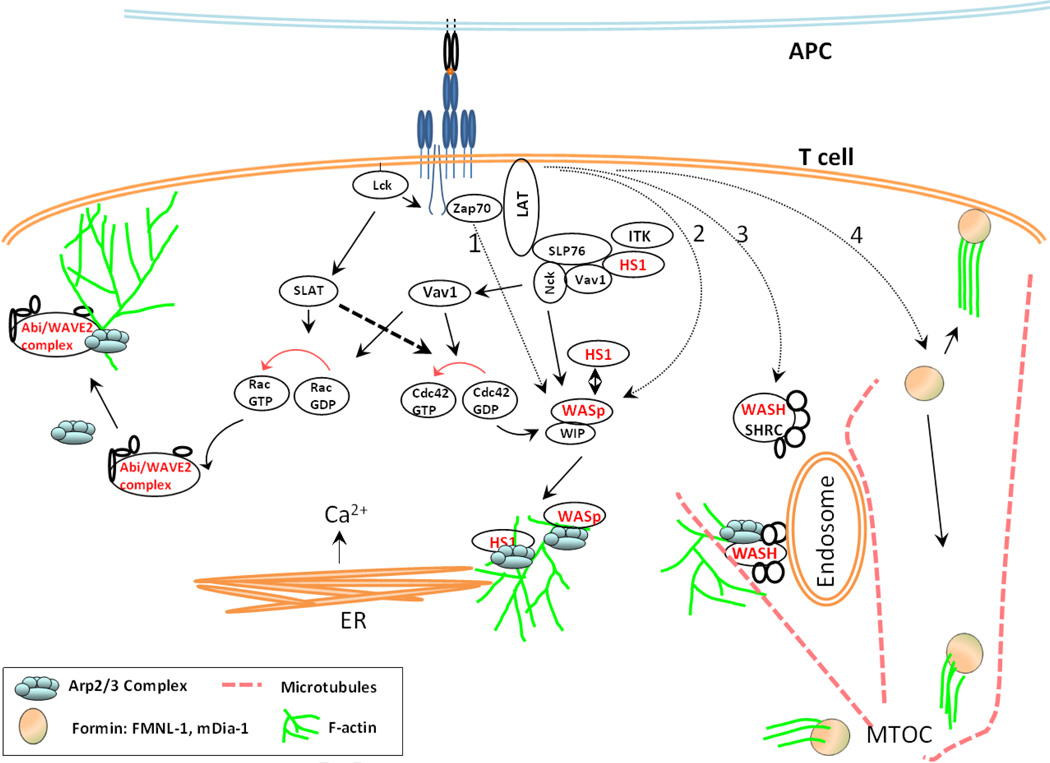
T cells constitute a crucial arm of the adaptive immune system and their optimal function is required for a healthy immune response. After the initial step of T cell-receptor (TCR) triggering by antigenic peptide complexes on antigen presenting cell (APC), the T cell exhibits extensive cytoskeletal remodeling. This cytoskeletal remodeling leads to the formation of an "immunological synapse" [1] characterized by regulated clustering, segregation and movement of receptors at the interface. Synapse formation regulates T cell activation and response to antigenic peptides and proceeds via feedback between actin cytoskeleton and TCR signaling. Actin polymerization participates in various events during the synapse formation, maturation, and eventually its disassembly. There is increasing knowledge about the actin effectors that couple TCR activation to actin rearrangements [2,3], and how defects in these effectors translate into impairment of T cell activation. In this review we aim to summarize and integrate parts of what is currently known about this feedback process. In addition, in light of recent advancements in our understanding of TCR triggering and translocation at the synapse, we speculate on the organizational and functional diversity of microfilament architecture in the T cell. This article is part of a Special Issue entitled: Reciprocal influences between cell cytoskeleton and membrane channels, receptors and transporters. Guest Editor: Jean Claude Hervé.
Keywords: Actin; Activation; Cytoskeleton; Lymphocytes; Myosin.
Copyright © 2013 Elsevier B.V. All rights reserved.
Figures
Similar articles
-
The calcium feedback loop and T cell activation: how cytoskeleton networks control intracellular calcium flux.Biochim Biophys Acta. 2014 Feb;1838(2):557-68. doi: 10.1016/j.bbamem.2013.07.009. Epub 2013 Jul 13. Biochim Biophys Acta. 2014. PMID: 23860253 Review.
-
The pivotal position of the actin cytoskeleton in the initiation and regulation of B cell receptor activation.Biochim Biophys Acta. 2014 Feb;1838(2):569-78. doi: 10.1016/j.bbamem.2013.07.016. Epub 2013 Jul 23. Biochim Biophys Acta. 2014. PMID: 23886914 Free PMC article. Review.
-
Regulation of T-cell receptor signaling by the actin cytoskeleton and poroelastic cytoplasm.Immunol Rev. 2013 Nov;256(1):148-59. doi: 10.1111/imr.12120. Immunol Rev. 2013. PMID: 24117819 Free PMC article. Review.
-
Recruitment of the actin-binding protein HIP-55 to the immunological synapse regulates T cell receptor signaling and endocytosis.J Biol Chem. 2004 Apr 9;279(15):15550-60. doi: 10.1074/jbc.M312659200. Epub 2004 Jan 16. J Biol Chem. 2004. PMID: 14729663
-
Multiple actin networks coordinate mechanotransduction at the immunological synapse.J Cell Biol. 2020 Feb 3;219(2):e201911058. doi: 10.1083/jcb.201911058. J Cell Biol. 2020. PMID: 31977034 Free PMC article.
Cited by
-
Dances with Membranes: Breakthroughs from Super-resolution Imaging.Curr Top Membr. 2015;75:59-123. doi: 10.1016/bs.ctm.2015.03.008. Epub 2015 Apr 15. Curr Top Membr. 2015. PMID: 26015281 Free PMC article. Review.
-
LPA suppresses T cell function by altering the cytoskeleton and disrupting immune synapse formation.Proc Natl Acad Sci U S A. 2022 Apr 12;119(15):e2118816119. doi: 10.1073/pnas.2118816119. Epub 2022 Apr 8. Proc Natl Acad Sci U S A. 2022. PMID: 35394866 Free PMC article.
-
Unraveling the Mechanobiology of the Immune System.Adv Healthc Mater. 2019 Feb;8(4):e1801332. doi: 10.1002/adhm.201801332. Epub 2019 Jan 7. Adv Healthc Mater. 2019. PMID: 30614636 Free PMC article. Review.
-
The PDL1-inducible GTPase Arl4d controls T effector function by limiting IL-2 production.Sci Rep. 2018 Oct 31;8(1):16123. doi: 10.1038/s41598-018-34522-4. Sci Rep. 2018. PMID: 30382149 Free PMC article.
-
Role of Actin Cytoskeleton Reorganization in Polarized Secretory Traffic at the Immunological Synapse.Front Cell Dev Biol. 2021 Feb 4;9:629097. doi: 10.3389/fcell.2021.629097. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33614660 Free PMC article. Review.
References
-
- Grakoui A, Bromley SK, Sumen C, Davis MM, Shaw AS, Allen PM, Dustin ML. The immunological synapse: a molecular machine controlling T cell activation. Science. 1999;285:221–227. - PubMed
-
- Burkhardt JK, Carrizosa E, Shaffer MH. The actin cytoskeleton in T cell activation. Annu Rev Immunol. 2008;26:233–259. - PubMed
-
- Reicher B, Barda-Saad M. Multiple pathways leading from the T-cell antigen receptor to the actin cytoskeleton network. FEBS Lett. 2010;584:4858–4864. - PubMed
-
- Irvine DJ, Purbhoo MA, Krogsgaard M, Davis MM. Direct observation of ligand recognition by T cells. Nature. 2002;419:845–849. - PubMed
-
- Thrasher AJ, Burns SO. WASP: a key immunological multitasker. Nature reviews. Immunology. 2010;10:182–192. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
