

Palmitoylation of the β4-subunit regulates surface expression of large conductance calcium-activated potassium channel splice variants
- PMID: 23504458
- PMCID: PMC3642354
- DOI: 10.1074/jbc.M113.461830
Palmitoylation of the β4-subunit regulates surface expression of large conductance calcium-activated potassium channel splice variants
Abstract
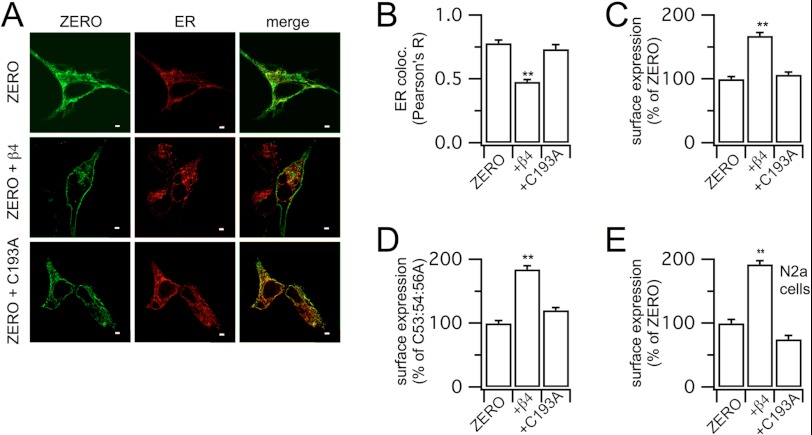
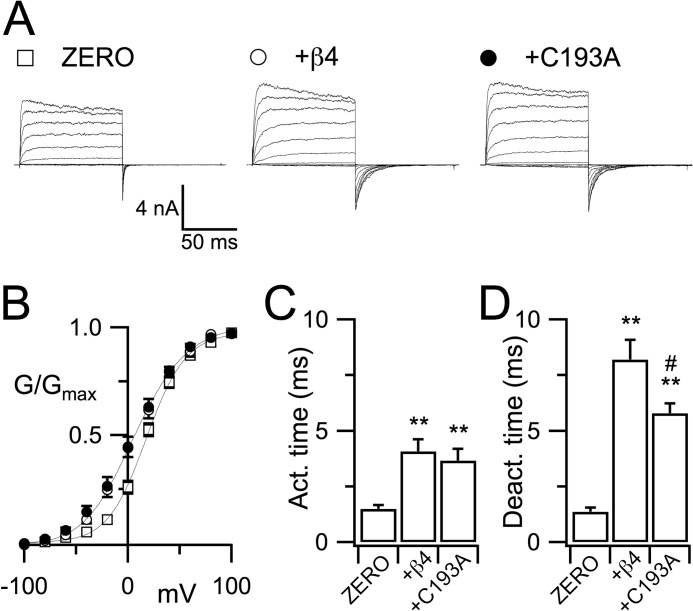
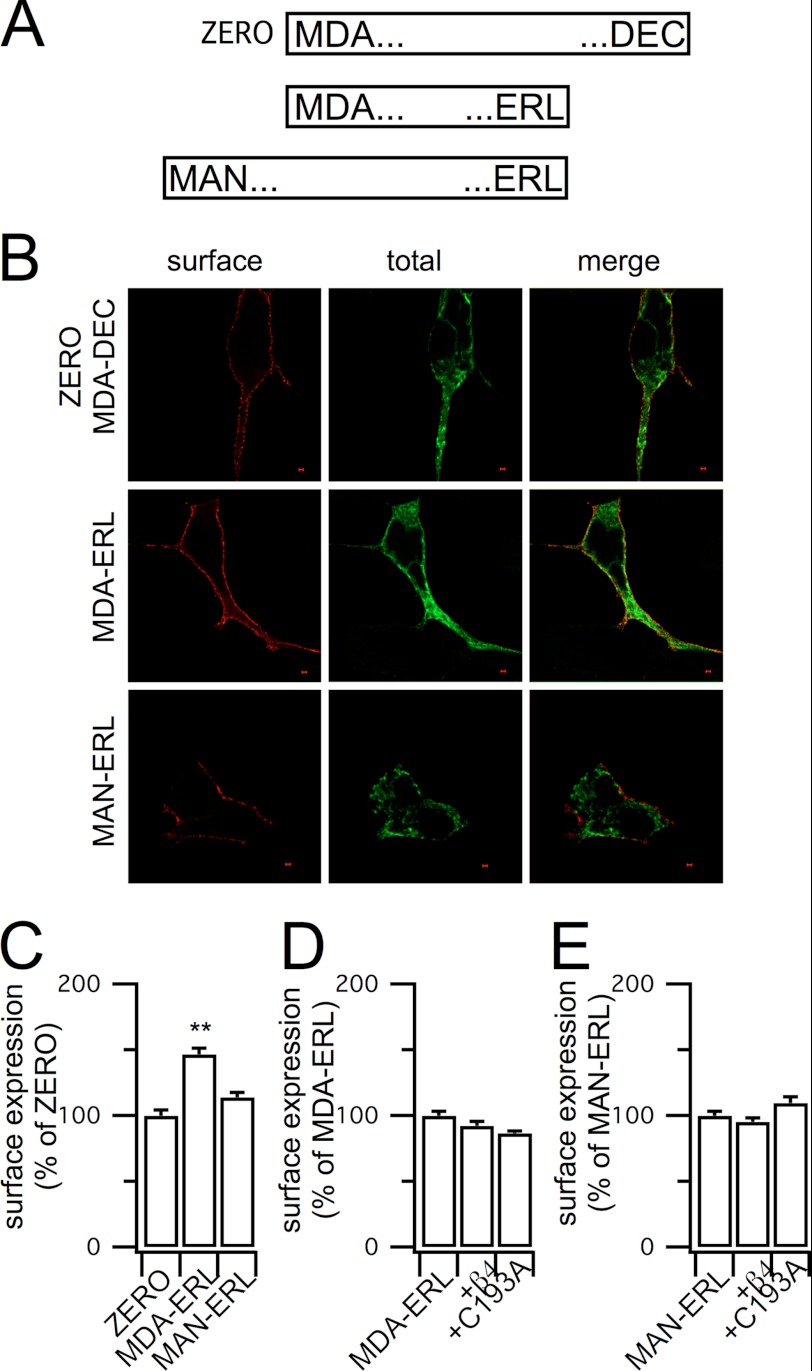
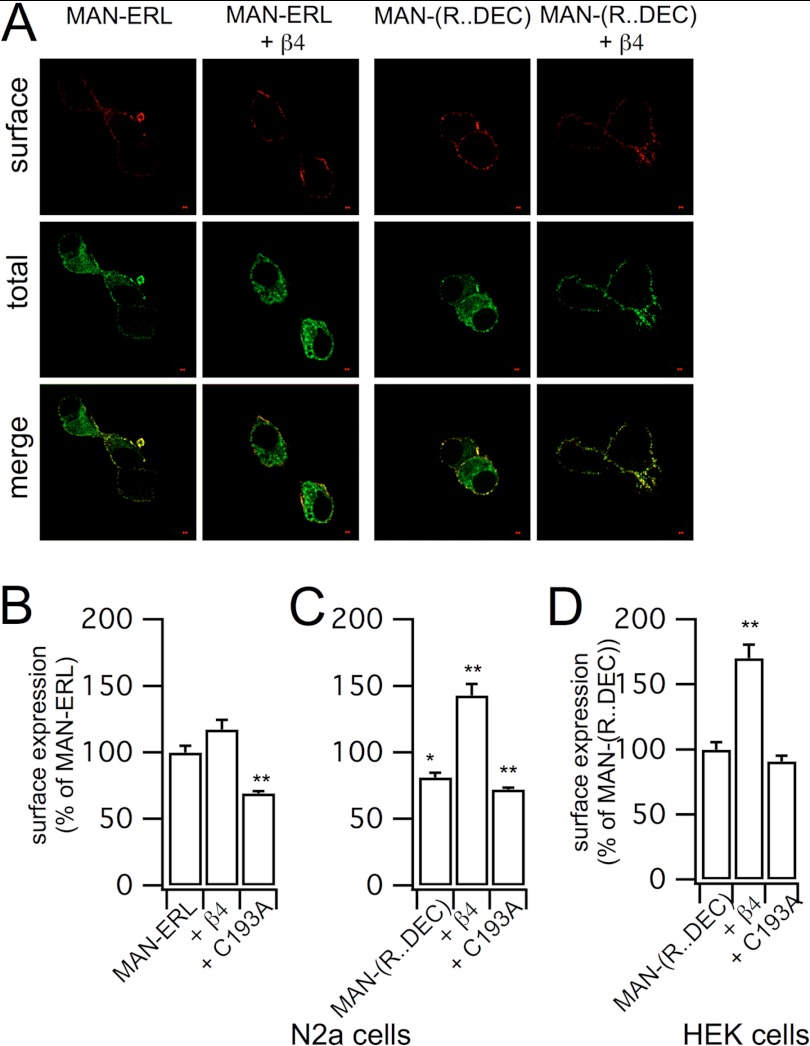
Regulatory β-subunits of large conductance calcium- and voltage-activated potassium (BK) channels play an important role in generating functional diversity and control of cell surface expression of the pore forming α-subunits. However, in contrast to α-subunits, the role of reversible post-translational modification of intracellular residues on β-subunit function is largely unknown. Here we demonstrate that the human β4-subunit is S-acylated (palmitoylated) on a juxtamembrane cysteine residue (Cys-193) in the intracellular C terminus of the regulatory β-subunit. β4-Subunit palmitoylation is important for cell surface expression and endoplasmic reticulum (ER) exit of the β4-subunit alone. Importantly, palmitoylated β4-subunits promote the ER exit and surface expression of the pore-forming α-subunit, whereas β4-subunits that cannot be palmitoylated do not increase ER exit or surface expression of α-subunits. Strikingly, however, this palmitoylation- and β4-dependent enhancement of α-subunit surface expression was only observed in α-subunits that contain a putative trafficking motif (… REVEDEC) at the very C terminus of the α-subunit. Engineering this trafficking motif to other C-terminal α-subunit splice variants results in α-subunits with reduced surface expression that can be rescued by palmitoylated, but not depalmitoylated, β4-subunits. Our data reveal a novel mechanism by which palmitoylated β4-subunit controls surface expression of BK channels through masking of a trafficking motif in the C terminus of the α-subunit. As palmitoylation is dynamic, this mechanism would allow precise control of specific splice variants to the cell surface. Our data provide new insights into how complex interplay between the repertoire of post-transcriptional and post-translational mechanisms controls cell surface expression of BK channels.
Keywords: Ion Channels; Membrane Transport; Potassium Channels; Protein Palmitoylation; S-Acylation; Signal Transduction.
Figures

Similar articles
-
Distinct domains of the β1-subunit cytosolic N terminus control surface expression and functional properties of large-conductance calcium-activated potassium (BK) channels.J Biol Chem. 2017 May 26;292(21):8694-8704. doi: 10.1074/jbc.M116.769505. Epub 2017 Apr 3. J Biol Chem. 2017. PMID: 28373283 Free PMC article.
-
Palmitoylation of the S0-S1 linker regulates cell surface expression of voltage- and calcium-activated potassium (BK) channels.J Biol Chem. 2010 Oct 22;285(43):33307-33314. doi: 10.1074/jbc.M110.153940. Epub 2010 Aug 6. J Biol Chem. 2010. PMID: 20693285 Free PMC article.
-
S-Acylation controls functional coupling of BK channel pore-forming α-subunits and β1-subunits.J Biol Chem. 2019 Aug 9;294(32):12066-12076. doi: 10.1074/jbc.RA119.009065. Epub 2019 Jun 18. J Biol Chem. 2019. PMID: 31213527 Free PMC article.
-
S-acylation dependent post-translational cross-talk regulates large conductance calcium- and voltage- activated potassium (BK) channels.Front Physiol. 2014 Aug 5;5:281. doi: 10.3389/fphys.2014.00281. eCollection 2014. Front Physiol. 2014. PMID: 25140154 Free PMC article. Review.
-
Regulation of BK Channels by Beta and Gamma Subunits.Annu Rev Physiol. 2019 Feb 10;81:113-137. doi: 10.1146/annurev-physiol-022516-034038. Annu Rev Physiol. 2019. PMID: 30742788 Free PMC article. Review.
Cited by
-
Current understanding of iberiotoxin-resistant BK channels in the nervous system.Front Physiol. 2014 Oct 9;5:382. doi: 10.3389/fphys.2014.00382. eCollection 2014. Front Physiol. 2014. PMID: 25346692 Free PMC article. Review.
-
Distinct domains of the β1-subunit cytosolic N terminus control surface expression and functional properties of large-conductance calcium-activated potassium (BK) channels.J Biol Chem. 2017 May 26;292(21):8694-8704. doi: 10.1074/jbc.M116.769505. Epub 2017 Apr 3. J Biol Chem. 2017. PMID: 28373283 Free PMC article.
-
Modulation of BK Channel Function by Auxiliary Beta and Gamma Subunits.Int Rev Neurobiol. 2016;128:51-90. doi: 10.1016/bs.irn.2016.03.015. Epub 2016 Apr 8. Int Rev Neurobiol. 2016. PMID: 27238261 Free PMC article. Review.
-
Regulatory effects of protein S-acylation on insulin secretion and insulin action.Open Biol. 2021 Mar;11(3):210017. doi: 10.1098/rsob.210017. Epub 2021 Mar 31. Open Biol. 2021. PMID: 33784857 Free PMC article. Review.
-
Mitochondrial BKCa channel.Front Physiol. 2015 Mar 31;6:104. doi: 10.3389/fphys.2015.00104. eCollection 2015. Front Physiol. 2015. PMID: 25873902 Free PMC article. Review.
References
-
- Orio P., Rojas P., Ferreira G., Latorre R. (2002) New disguises for an old channel: MaxiK channel β-subunits. News Physiol. Sci. 17, 156–161 - PubMed
-
- Brenner R., Peréz G. J., Bonev A. D., Eckman D. M., Kosek J. C., Wiler S. W., Patterson A. J., Nelson M. T., Aldrich R. W. (2000) Vasoregulation by the β1 subunit of the calcium-activated potassium channel. Nature 407, 870–876 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
