

Review
doi: 10.1016/j.ceb.2013.02.017.
Epub 2013 Mar 13.
5-Hydroxymethylcytosine: generation, fate, and genomic distribution
Affiliations
- PMID: 23498661
- PMCID: PMC4060438
- DOI: 10.1016/j.ceb.2013.02.017
Item in Clipboard
Review
5-Hydroxymethylcytosine: generation, fate, and genomic distribution
Curr Opin Cell Biol.
2013 Jun.
Abstract
5-Methylcytosine (5mC) can be converted to 5-hydroxymethylcytosine (5hmC) in mammalian cells by the ten-eleven translocation (Tet) family of dioxygenases. While 5mC has been extensively studied, we have just started to understand the distribution and function of 5hmC in mammalian genomes. Despite the fact that this new epigenetic mark has only been discovered three years ago, exciting progress has been made in understanding its generation, fate, and genomic distribution. In this review we discuss these progresses as well as the recent advance in the single-base resolution mapping of 5hmC.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
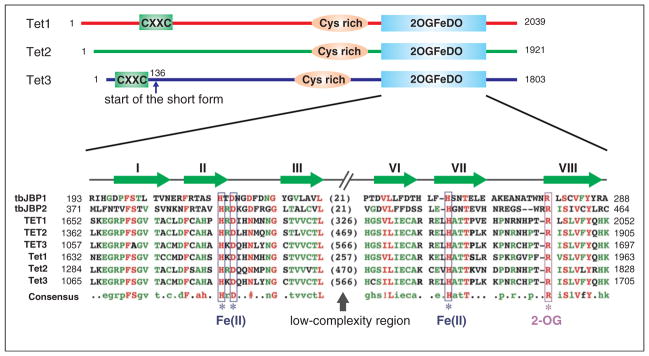
Schematic diagrams of the Tet proteins. Three conserved domains are indicated in mouse Tet proteins, including CXXC zinc finger, cysteine-rich region (Cys-rich), and the double-stranded β-helix (DSBH) fold of the 2OG-dependent and Fe(II)-dependent dioxygenase domain (2OGFeDO). Note that Tet3 has a shorter form, which starts at amino acid 136, that does not contain the CXXC domain. For the 2OGFeDO domain, a multiple sequence alignment of selected JBP/Tet family proteins is shown. Sequences used in the alignment include the Trypanosoma brucei JBP1 (Q9U6M3) and JBP2 (Q57X81); human TET1 (NP_085128), TET2 (NP_001120680), and TET3 (ADU77107); and mouse Tet1 (NP_001240786), Tet2 (ACY38292), and Tet3 (ADR57137). Predicted Fe(II) and 2OG-binding sites are indicated, and the conserved strands that constitute the DSBH fold are shown above the multiple sequence alignment. Numbers represent the amino acid numbers.

Proposed DNA demethylation pathways that involve 5hmC. DNA methylation (5mC) is established and maintained by DNA methyltransferases (DNMTs). In mammals, 5mC can be oxidized by the Tet proteins to generate 5hmC. 5hmC is recognized poorly by Dnmt1 and can be diluted during DNA replication. 5hmC can also be further oxidized by Tet proteins to produce 5fC and 5caC. Alternatively, 5hmC may be deaminated by AID/APOBECs to become 5hmU. 5fC, 5caC, and 5hmU can be excised from DNA by glycosylases. In addition, DNMT3A and DNMT3B may directly dehydroxymethylate 5hmC to generate unmodified C. Note that solid lines represent processes with strong evidence, while the dashed lines indicate processes which need to be further confirmed controversial process.
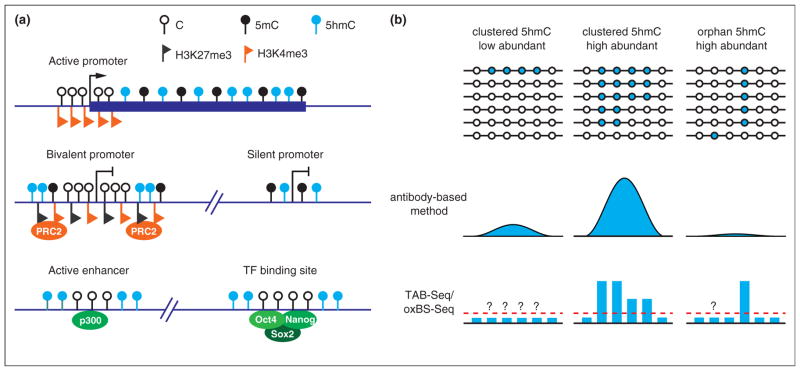
Genomic distribution of 5hmC. 1. Schematic diagram illustrating the 5hmC distribution in the genome of mouse ESCs. 5hmC is preferentially enriched at gene bodies of actively transcribed genes, bivalent and silent promoters, as well as active enhancers and a cohort of pluripotency transcription factor binding sites. 2. Comparison between the antibody-based 5hmC profiling methods and the single-base resolution methods. Antibody-based methods are not sensitive enough to detect all orphan 5hmCs, and thus may not be able to detect low 5hmC-density regions. In contrast, the single-base resolution methods can quantitatively determine the levels of orphan 5hmCs. However, the single-base resolution methods still cannot achieve low detection limitation (red dashed line) with normal sequencing depth. Thus, 5hmCs with low abundance in a population of cells may not be confidently identified.
Similar articles
-
Charting oxidized methylcytosines at base resolution.Nat Struct Mol Biol. 2015 Sep;22(9):656-61. doi: 10.1038/nsmb.3071. Nat Struct Mol Biol. 2015. PMID: 26333715 Free PMC article.
-
Tet family of 5-methylcytosine dioxygenases in mammalian development.J Hum Genet. 2013 Jul;58(7):421-7. doi: 10.1038/jhg.2013.63. Epub 2013 May 30. J Hum Genet. 2013. PMID: 23719188 Free PMC article. Review.
-
High-Resolution Analysis of 5-Hydroxymethylcytosine by TET-Assisted Bisulfite Sequencing.Methods Mol Biol. 2021;2198:321-331. doi: 10.1007/978-1-0716-0876-0_25. Methods Mol Biol. 2021. PMID: 32822042
-
Genomic distribution and possible functions of DNA hydroxymethylation in the brain.Genomics. 2014 Nov;104(5):341-6. doi: 10.1016/j.ygeno.2014.08.020. Epub 2014 Sep 7. Genomics. 2014. PMID: 25205307 Review.
-
A TET homologue protein from Coprinopsis cinerea (CcTET) that biochemically converts 5-methylcytosine to 5-hydroxymethylcytosine, 5-formylcytosine, and 5-carboxylcytosine.J Am Chem Soc. 2014 Apr 2;136(13):4801-4. doi: 10.1021/ja500979k. Epub 2014 Mar 25. J Am Chem Soc. 2014. PMID: 24655109 Free PMC article.
Cited by
-
The methylcytosine dioxygenase Tet2 promotes DNA demethylation and activation of cytokine gene expression in T cells.Immunity. 2015 Apr 21;42(4):613-26. doi: 10.1016/j.immuni.2015.03.005. Epub 2015 Apr 7. Immunity. 2015. PMID: 25862091 Free PMC article.
-
Loss of Tet1-Associated 5-Hydroxymethylcytosine Is Concomitant with Aberrant Promoter Hypermethylation in Liver Cancer.Cancer Res. 2016 May 15;76(10):3097-108. doi: 10.1158/0008-5472.CAN-15-1910. Epub 2016 Apr 6. Cancer Res. 2016. PMID: 27197233 Free PMC article.
-
UHRF2 regulates cell cycle, epigenetics and gene expression to control the timing of retinal progenitor and ganglion cell differentiation.Development. 2022 Mar 15;149(6):dev195644. doi: 10.1242/dev.195644. Epub 2022 Mar 14. Development. 2022. PMID: 35285483 Free PMC article.
-
Bisulfite-free, single base-resolution analysis of 5-hydroxymethylcytosine in genomic DNA by chemical-mediated mismatch.Chem Sci. 2018 Oct 11;10(2):447-452. doi: 10.1039/c8sc04272a. eCollection 2019 Jan 14. Chem Sci. 2018. PMID: 30746092 Free PMC article.
-
Single-locus enrichment without amplification for sequencing and direct detection of epigenetic modifications.Mol Genet Genomics. 2016 Jun;291(3):1491-504. doi: 10.1007/s00438-016-1167-2. Epub 2016 Jan 29. Mol Genet Genomics. 2016. PMID: 26825750
References
-
- He YF, Li BZ, Li Z, Liu P, Wang Y, Tang Q, Ding J, Jia Y, Chen Z, Li L, et al. Tet-mediated formation of 5-carboxylcytosine and its excision by TDG in mammalian DNA. Science. 2011;333:1303–1307. This paper demonstrates that 5mC and 5hmC can be oxidized to 5caC by Tet proteins. The authors also show that 5caC can be specifically recognized and excised from DNA by TDG. - PMC - PubMed
-
- Ito S, Shen L, Dai Q, Wu SC, Collins LB, Swenberg JA, He C, Zhang Y. Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science. 2011;333:1300–1303. This paper demonstrates that Tet dioxygenases can convert 5mC and 5hmC to both 5fC and 5caC in vitro and in vivo in an enzymatic activity-dependent manner. The authors also quantified the absolute amounts of all these cytosine derivatives in various mouse cell lines and organs. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
