

Plasmin-mediated activation of pandemic H1N1 influenza virus hemagglutinin is independent of the viral neuraminidase
- PMID: 23449787
- PMCID: PMC3624321
- DOI: 10.1128/JVI.00210-13
Plasmin-mediated activation of pandemic H1N1 influenza virus hemagglutinin is independent of the viral neuraminidase
Abstract
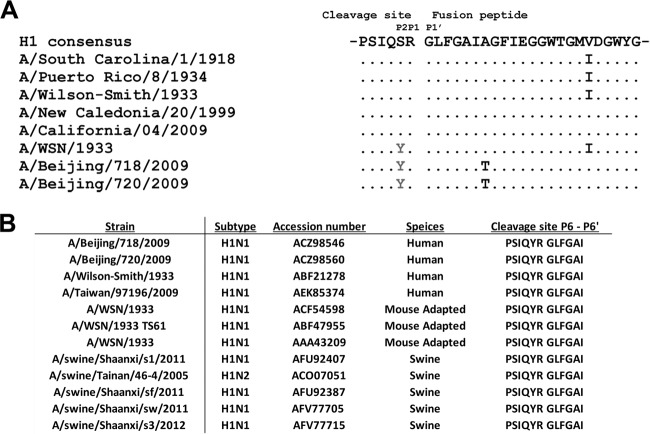
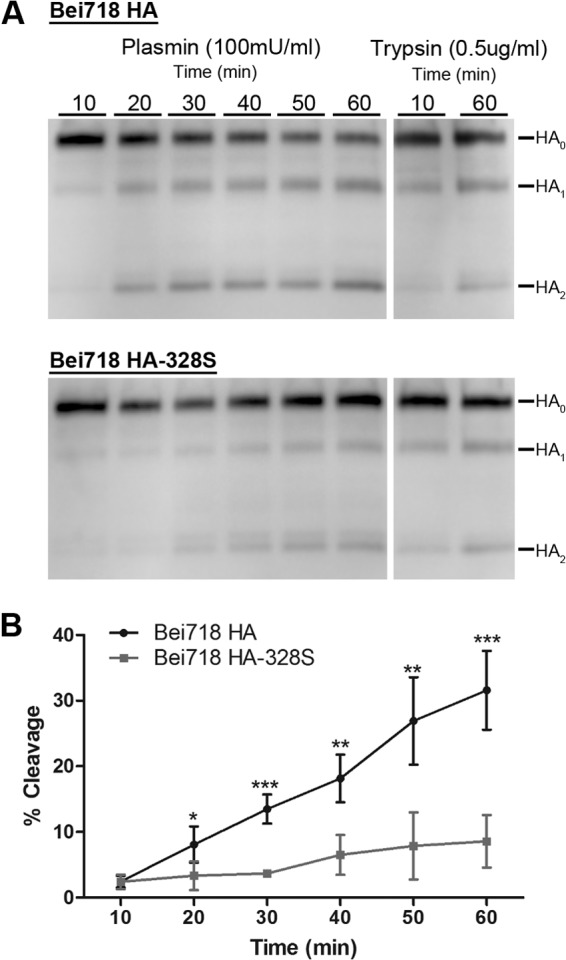
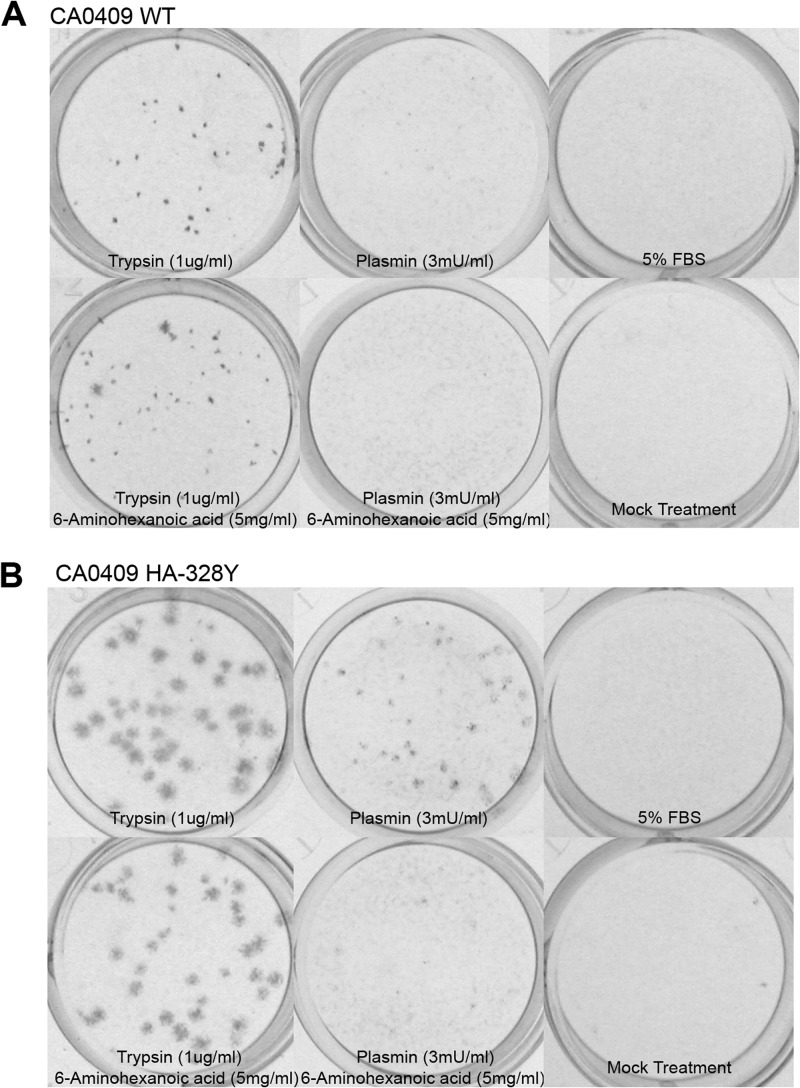
Influenza virus is well recognized to modulate host tropism and pathogenesis based on mutations in the proteolytic cleavage site of the viral hemagglutinin (HA), which activates HA and exposes the fusion peptide for membrane fusion. Instead of the conventional trypsin-mediated cleavage event, modification of the cleavage site allows extended use of host cell proteases and enhanced spread in vivo. For H1N1 influenza viruses, the mouse-adapted A/WSN/33 strain is known to replicate in the brain based on recruitment of plasminogen by the viral neuraminidase (NA), as well as a Ser-Tyr substitution at the P2 position of the HA cleavage site. Here, we show that an equivalent Ser-Tyr substitution has occurred in the HA of naturally occurring human H1N1 influenza viruses. We characterize one of these viruses (A/Beijing/718/2009), as well as the prototype A/California/04/2009 with a Ser-Tyr substitution in the cleavage site, and show that these HAs are preferentially cleaved by plasmin. Importantly, cleavage activation by plasmin/plasminogen was independent of the viral NA, suggesting a novel mechanism for HA cleavage activation. We show that the viral HA itself can recruit plasminogen for HA cleavage. We further show that cellular factors, as well as streptokinase from bacteria commonly coinfecting the respiratory tract of influenza patients, can be a source of activated plasminogen for plasmin-mediated cleavage of influenza virus HAs that contain a Ser-Tyr substitution in the cleavage site.
Figures
Similar articles
-
Modifications to the hemagglutinin cleavage site control the virulence of a neurotropic H1N1 influenza virus.J Virol. 2010 Sep;84(17):8683-90. doi: 10.1128/JVI.00797-10. Epub 2010 Jun 16. J Virol. 2010. PMID: 20554779 Free PMC article.
-
Functional balance of the hemagglutinin and neuraminidase activities accompanies the emergence of the 2009 H1N1 influenza pandemic.J Virol. 2012 Sep;86(17):9221-32. doi: 10.1128/JVI.00697-12. Epub 2012 Jun 20. J Virol. 2012. PMID: 22718832 Free PMC article.
-
Positive selection on hemagglutinin and neuraminidase genes of H1N1 influenza viruses.Virol J. 2011 Apr 21;8:183. doi: 10.1186/1743-422X-8-183. Virol J. 2011. PMID: 21507270 Free PMC article.
-
[Advances in the structure and function of pandemic A/H1N1/2009 influenza virus HA protein].Bing Du Xue Bao. 2012 Jun;28(4):444-52. Bing Du Xue Bao. 2012. PMID: 22978172 Review. Chinese.
-
Competitive Cooperation of Hemagglutinin and Neuraminidase during Influenza A Virus Entry.Viruses. 2019 May 20;11(5):458. doi: 10.3390/v11050458. Viruses. 2019. PMID: 31137516 Free PMC article. Review.
Cited by
-
Necrotic Response to Low Pathogenic H9N2 Influenza Virus in Chicken Hepatoma Cells.Jundishapur J Microbiol. 2015 Jan 25;8(1):e13770. doi: 10.5812/jjm.13770. eCollection 2015 Jan. Jundishapur J Microbiol. 2015. PMID: 25789133 Free PMC article.
-
Human matriptase/ST 14 proteolytically cleaves H7N9 hemagglutinin and facilitates the activation of influenza A/Shanghai/2/2013 virus in cell culture.Influenza Other Respir Viruses. 2020 Mar;14(2):189-195. doi: 10.1111/irv.12707. Epub 2019 Dec 9. Influenza Other Respir Viruses. 2020. PMID: 31820577 Free PMC article.
-
Host cell proteases: Critical determinants of coronavirus tropism and pathogenesis.Virus Res. 2015 Apr 16;202:120-34. doi: 10.1016/j.virusres.2014.11.021. Epub 2014 Nov 22. Virus Res. 2015. PMID: 25445340 Free PMC article. Review.
-
Influenza A(H1N1)pdm09 Virus Aggravates Pathology of Blood Vessels in Wistar Rats with Premorbid Acute Cardiomyopathy.Viruses. 2023 May 4;15(5):1114. doi: 10.3390/v15051114. Viruses. 2023. PMID: 37243200 Free PMC article.
-
Equine and Canine Influenza H3N8 Viruses Show Minimal Biological Differences Despite Phylogenetic Divergence.J Virol. 2015 Jul;89(13):6860-73. doi: 10.1128/JVI.00521-15. Epub 2015 Apr 22. J Virol. 2015. PMID: 25903329 Free PMC article.
References
-
- Cox NJ, Subbarao K. 2000. Global epidemiology of influenza: past and present. Annu. Rev. Med. 51:407–421 - PubMed
-
- Wiley DC, Skehel JJ. 1987. The structure and function of the hemagglutinin membrane glycoprotein of influenza virus. Annu. Rev. Biochem. 56:365–394 - PubMed
-
- Klenk H-D, Garten W. 1994. Activation cleavage of viral spike proteins by host proteases, p 241–280 In Wimmer E. (ed), Cellular receptors for animal viruses. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
-
- Klenk HD, Garten W. 1994. Host cell proteases controlling virus pathogenicity. Trends Microbiol. 2:39–43 - PubMed
-
- Choi SY, Bertram S, Glowacka I, Park YW, Pohlmann S. 2009. Type II transmembrane serine proteases in cancer and viral infections. Trends Mol. Med. 15:303–312 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
