

The maternal to zygotic transition in mammals
- PMID: 23352575
- PMCID: PMC3669654
- DOI: 10.1016/j.mam.2013.01.003
The maternal to zygotic transition in mammals
Abstract
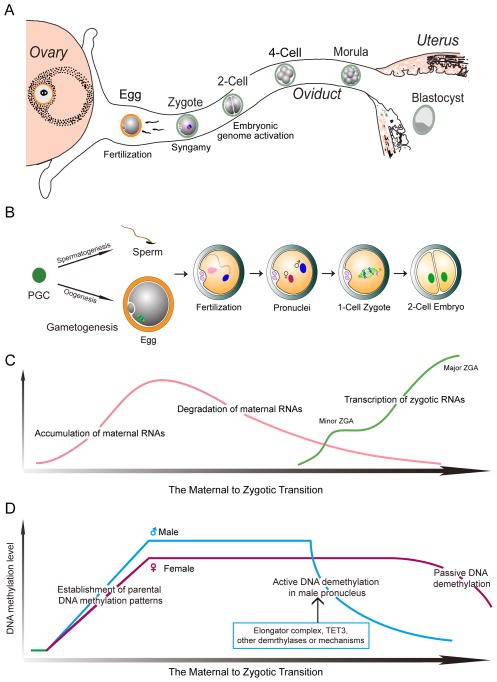
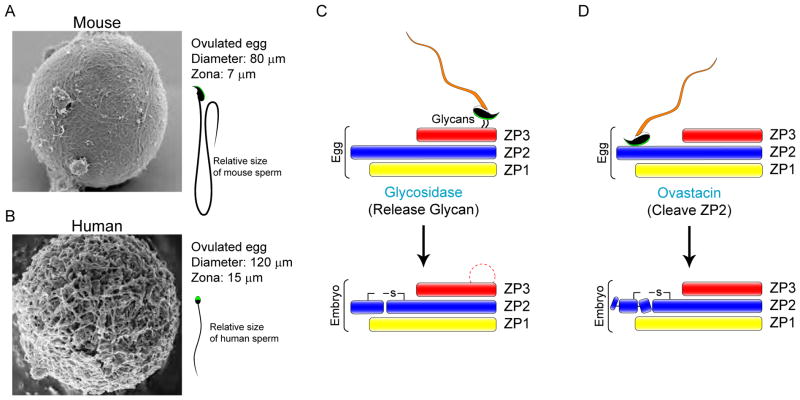
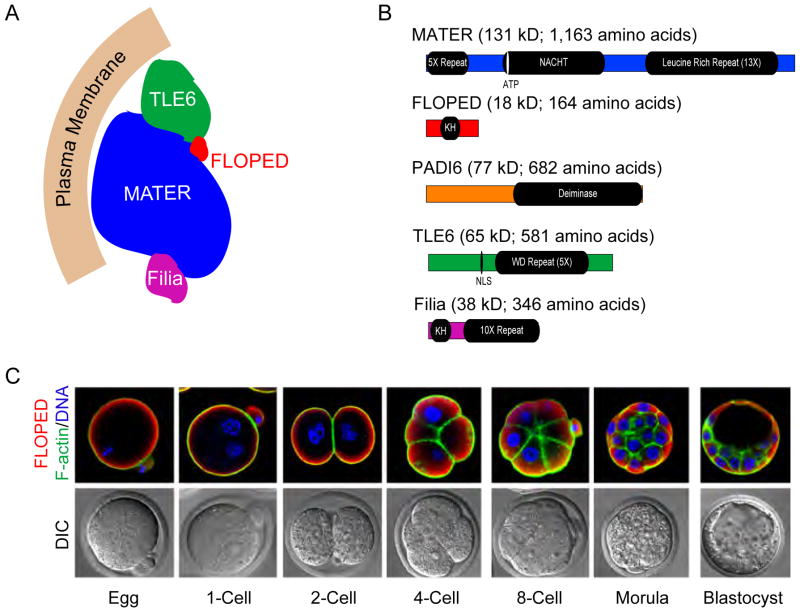
Prior to activation of the embryonic genome, the initiating events of mammalian development are under maternal control and include fertilization, the block to polyspermy and processing sperm DNA. Following gamete union, the transcriptionally inert sperm DNA is repackaged into the male pronucleus which fuses with the female pronucleus to form a 1-cell zygote. Embryonic transcription begins during the maternal to zygotic transfer of control in directing development. This transition occurs at species-specific times after one or several rounds of blastomere cleavage and is essential for normal development. However, even after activation of the embryonic genome, successful development relies on stored maternal components without which embryos fail to progress beyond initial cell divisions. Better understanding of the molecular basis of maternal to zygotic transition including fertilization, the activation of the embryonic genome and cleavage-stage development will provide insight into early human development that should translate into clinical applications for regenerative medicine and assisted reproductive technologies.
Keywords: 5-carboxylcytosine; 5-formylcytosine; 5caC; 5fC; ART; DMR; DNA methylation; DNA methyltransferases; DNMTs; Gamete recognition; Histone modification; ICR; MZT; Maternal effect genes; Maternal to zygotic transition (MZT); Mouse fertilization; Piwi-interacting RNA; RNA interference; RNAi; SCMC; SCNT; Subcortical maternal complex (SCMC); TDG; TRC; ZGA; Zygotic genome activation (ZGA); assisted reproductive technology; differentially methylated regions; double strand RNA; dsRNA; endo-siRNA; endo-small interfering RNA; imprinting control regions; maternal to zygotic transition; miRNA; microRNA; piRNA; somatic cell nuclear transfer; subcortical maternal complex; thymine-DNA glycosylase; transcription required complex; zygotic genome activation.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
A story of birth and death: mRNA translation and clearance at the onset of maternal-to-zygotic transition in mammals†.Biol Reprod. 2019 Sep 1;101(3):579-590. doi: 10.1093/biolre/ioz012. Biol Reprod. 2019. PMID: 30715134 Review.
-
Zygotic genome activation in the chicken: a comparative review.Cell Mol Life Sci. 2020 May;77(10):1879-1891. doi: 10.1007/s00018-019-03360-6. Epub 2019 Nov 15. Cell Mol Life Sci. 2020. PMID: 31728579 Free PMC article. Review.
-
Post-translational regulation of the maternal-to-zygotic transition.Cell Mol Life Sci. 2018 May;75(10):1707-1722. doi: 10.1007/s00018-018-2750-y. Epub 2018 Feb 9. Cell Mol Life Sci. 2018. PMID: 29427077 Free PMC article. Review.
-
Dynamics of TET family expression in porcine preimplantation embryos is related to zygotic genome activation and required for the maintenance of NANOG.Dev Biol. 2014 Feb 1;386(1):86-95. doi: 10.1016/j.ydbio.2013.11.024. Epub 2013 Dec 4. Dev Biol. 2014. PMID: 24315853
-
Cell Cycle Remodeling and Zygotic Gene Activation at the Midblastula Transition.Adv Exp Med Biol. 2017;953:441-487. doi: 10.1007/978-3-319-46095-6_9. Adv Exp Med Biol. 2017. PMID: 27975277 Review.
Cited by
-
The periconceptional environment and cardiovascular disease: does in vitro embryo culture and transfer influence cardiovascular development and health?Nutrients. 2015 Feb 18;7(3):1378-425. doi: 10.3390/nu7031378. Nutrients. 2015. PMID: 25699984 Free PMC article. Review.
-
The H3.3 chaperone Hira complex orchestrates oocyte developmental competence.Development. 2022 Mar 1;149(5):dev200044. doi: 10.1242/dev.200044. Epub 2022 Feb 28. Development. 2022. PMID: 35112132 Free PMC article.
-
Free fatty acid treatment of mouse preimplantation embryos demonstrates contrasting effects of palmitic acid and oleic acid on autophagy.Am J Physiol Cell Physiol. 2022 May 1;322(5):C833-C848. doi: 10.1152/ajpcell.00414.2021. Epub 2022 Mar 23. Am J Physiol Cell Physiol. 2022. PMID: 35319901 Free PMC article.
-
Arginine Regulates Zygotic Genome Activation in Porcine Embryos Under Nutrition Restriction.Front Vet Sci. 2022 Jun 23;9:921406. doi: 10.3389/fvets.2022.921406. eCollection 2022. Front Vet Sci. 2022. PMID: 35812864 Free PMC article.
-
Dppa2 and Dppa4 directly regulate the Dux-driven zygotic transcriptional program.Genes Dev. 2019 Feb 1;33(3-4):194-208. doi: 10.1101/gad.321174.118. Epub 2019 Jan 28. Genes Dev. 2019. PMID: 30692203 Free PMC article.
References
-
- Aapola U, Kawasaki K, Scott HS, Ollila J, Vihinen M, Heino M, Shintani A, Minoshima S, Krohn K, Antonarakis SE, Shimizu N, Kudoh J, Peterson P. Isolation and initial characterization of a novel zinc finger gene, DNMT3L, on 21q22.3, related to the cytosine-5-methyltransferase 3 gene family. Genomics. 2000;65 (3):293–298. - PubMed
-
- Abdalla H, Yoshizawa Y, Hochi S. Active demethylation of paternal genome in mammalian zygotes. J Reprod Dev. 2009;55 (4):356–360. - PubMed
-
- Adenot PG, Mercier Y, Renard JP, Thompson EM. Differential H4 acetylation of paternal and maternal chromatin precedes DNA replication and differential transcriptional activity in pronuclei of 1-cell mouse embryos. Development. 1997;124 (22):4615–4625. - PubMed
-
- Aoki F, Worrad DM, Schultz RM. Regulation of transcriptional activity during the first and second cell cycles in the preimplantation mouse embryo. Dev Biol. 1997;181 (2):296–307. - PubMed
-
- Arney KL, Bao S, Bannister AJ, Kouzarides T, Surani MA. Histone methylation defines epigenetic asymmetry in the mouse zygote. Int J Dev Biol. 2002;46 (3):317–320. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
