

Energy metabolism regulates clathrin adaptors at the trans-Golgi network and endosomes
- PMID: 23345590
- PMCID: PMC3596253
- DOI: 10.1091/mbc.E12-10-0750
Energy metabolism regulates clathrin adaptors at the trans-Golgi network and endosomes
Abstract
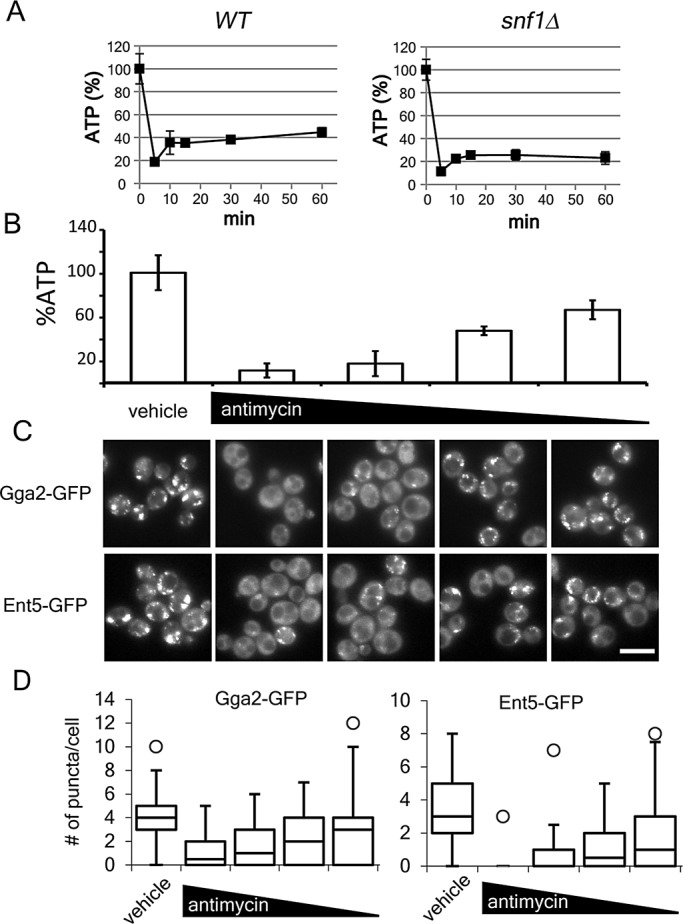
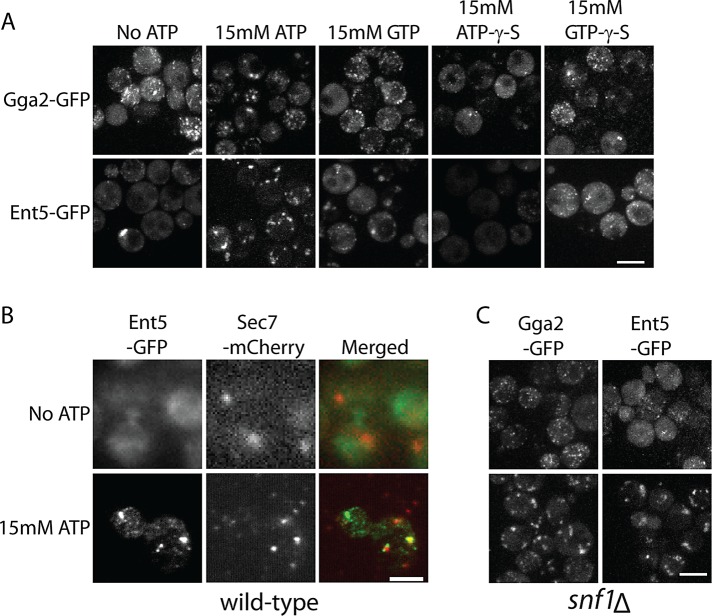
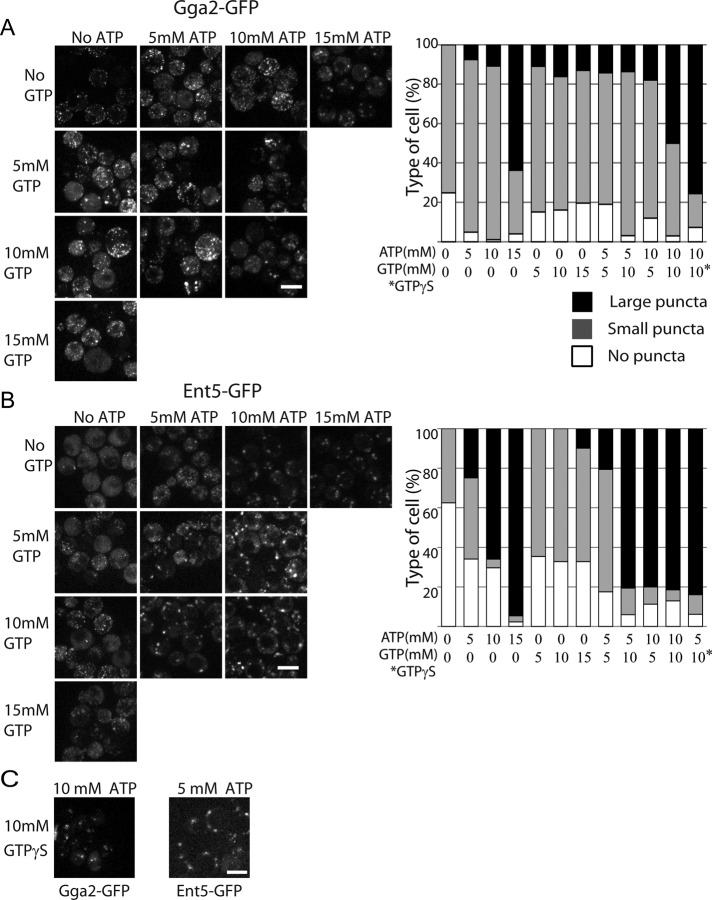
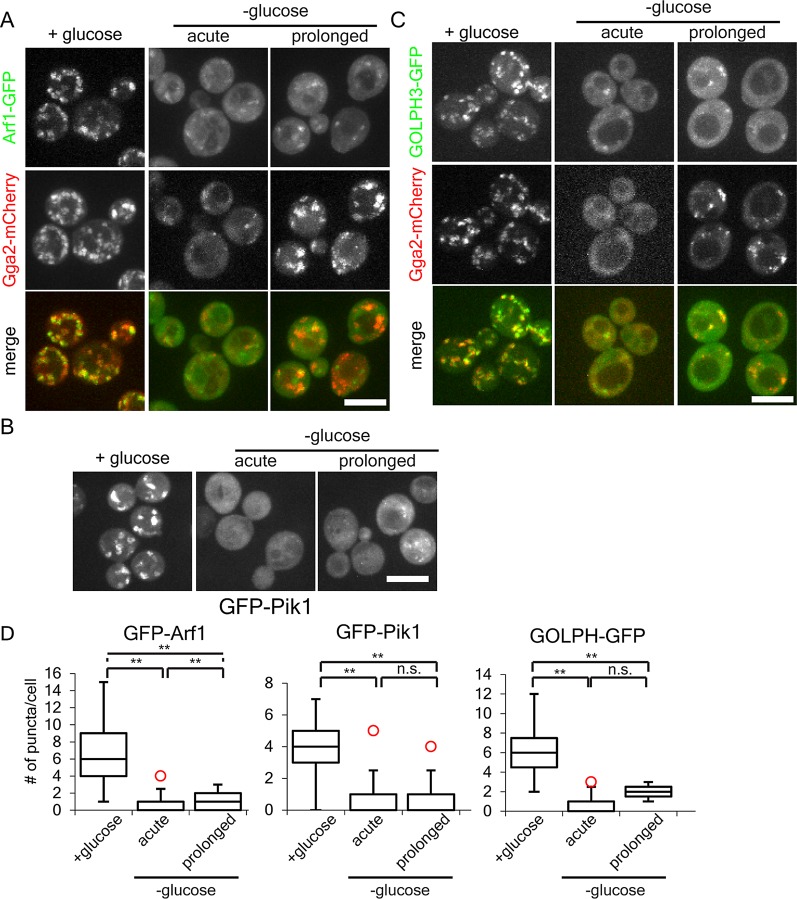
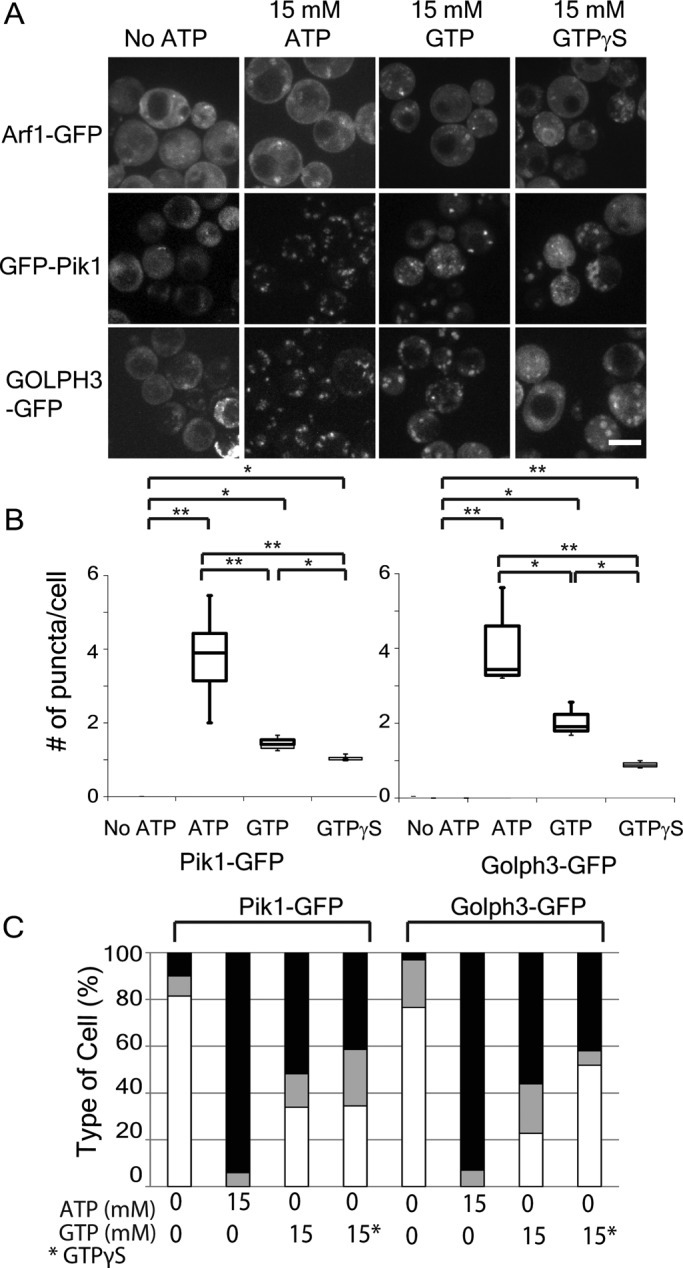
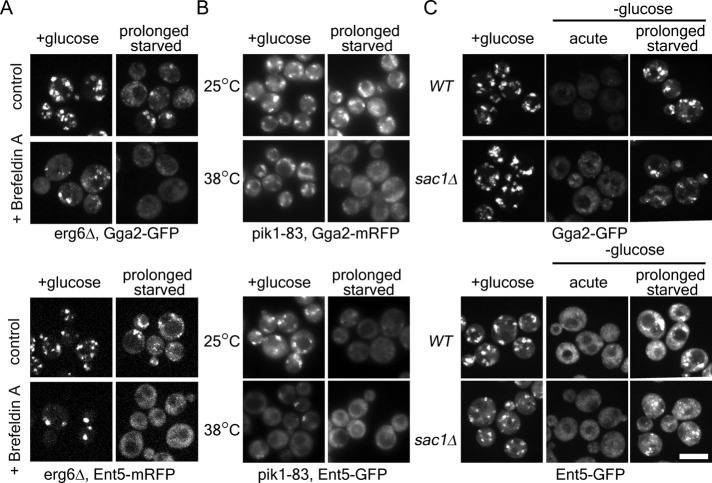
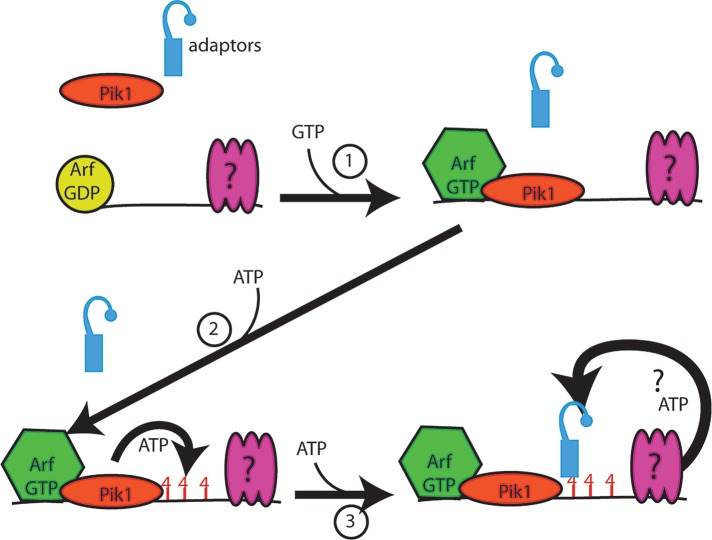
Glucose is a master regulator of cell behavior in the yeast Saccharomyces cerevisiae. It acts as both a metabolic substrate and a potent regulator of intracellular signaling cascades. Glucose starvation induces the transient delocalization and then partial relocalization of clathrin adaptors at the trans-Golgi network and endosomes. Although these localization responses are known to depend on the protein kinase A (PKA) signaling pathway, the molecular mechanism of this regulation is unknown. Here we demonstrate that PKA and the AMP-regulated kinase regulate adaptor localization through changes in energy metabolism. We show that genetic and chemical manipulation of intracellular ATP levels cause corresponding changes in adaptor localization. In permeabilized cells, exogenous ATP is sufficient to induce adaptor localization. Furthermore, we reveal distinct energy-dependent steps in adaptor localization: a step that requires the ADP-ribosylation factor ARF, an ATP-dependent step that requires the phosphatidyl-inositol-4 kinase Pik1, and third ATP-dependent step for which we provide evidence but for which the mechanism is unknown. We propose that these energy-dependent mechanisms precisely synchronize membrane traffic with overall proliferation rates and contribute a crucial aspect of energy conservation during acute glucose starvation.
Figures
Similar articles
-
Glucose regulates clathrin adaptors at the trans-Golgi network and endosomes.Mol Biol Cell. 2011 Oct;22(19):3671-83. doi: 10.1091/mbc.E11-04-0309. Epub 2011 Aug 10. Mol Biol Cell. 2011. PMID: 21832155 Free PMC article.
-
Conserved role for Gga proteins in phosphatidylinositol 4-kinase localization to the trans-Golgi network.Proc Natl Acad Sci U S A. 2017 Mar 28;114(13):3433-3438. doi: 10.1073/pnas.1615163114. Epub 2017 Mar 13. Proc Natl Acad Sci U S A. 2017. PMID: 28289207 Free PMC article.
-
Phosphoinositide-mediated clathrin adaptor progression at the trans-Golgi network.Nat Cell Biol. 2012 Feb 19;14(3):239-48. doi: 10.1038/ncb2427. Nat Cell Biol. 2012. PMID: 22344030 Free PMC article.
-
New directions for the clathrin adaptor AP-1 in cell biology and human disease.Curr Opin Cell Biol. 2022 Jun;76:102079. doi: 10.1016/j.ceb.2022.102079. Epub 2022 Apr 13. Curr Opin Cell Biol. 2022. PMID: 35429729 Free PMC article. Review.
-
GGA proteins: new players in the sorting game.J Cell Sci. 2001 Oct;114(Pt 19):3413-8. doi: 10.1242/jcs.114.19.3413. J Cell Sci. 2001. PMID: 11682601 Review.
Cited by
-
Metabolic regulation through the endosomal system.Traffic. 2019 Aug;20(8):552-570. doi: 10.1111/tra.12670. Epub 2019 Jun 24. Traffic. 2019. PMID: 31177593 Free PMC article. Review.
-
AMP-Activated Protein Kinase Regulates the Cell Surface Proteome and Integrin Membrane Traffic.PLoS One. 2015 May 26;10(5):e0128013. doi: 10.1371/journal.pone.0128013. eCollection 2015. PLoS One. 2015. PMID: 26010094 Free PMC article.
-
Caloric restriction controls stationary phase survival through Protein Kinase A (PKA) and cytosolic pH.Aging Cell. 2019 Jun;18(3):e12921. doi: 10.1111/acel.12921. Epub 2019 Feb 20. Aging Cell. 2019. PMID: 30790427 Free PMC article.
-
Conserved Autophagy Pathway Contributes to Stress Tolerance and Virulence and Differentially Controls Autophagic Flux Upon Nutrient Starvation in Cryptococcus neoformans.Front Microbiol. 2019 Nov 26;10:2690. doi: 10.3389/fmicb.2019.02690. eCollection 2019. Front Microbiol. 2019. PMID: 32038502 Free PMC article.
-
Rapid glucose depletion immobilizes active myosin V on stabilized actin cables.Curr Biol. 2014 Oct 20;24(20):2471-9. doi: 10.1016/j.cub.2014.09.017. Epub 2014 Oct 9. Curr Biol. 2014. PMID: 25308080 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
