

Effects on vesicular transport pathways at the late endosome in cells with limited very long-chain fatty acids
- PMID: 23325927
- PMCID: PMC3617957
- DOI: 10.1194/jlr.M034678
Effects on vesicular transport pathways at the late endosome in cells with limited very long-chain fatty acids
Abstract
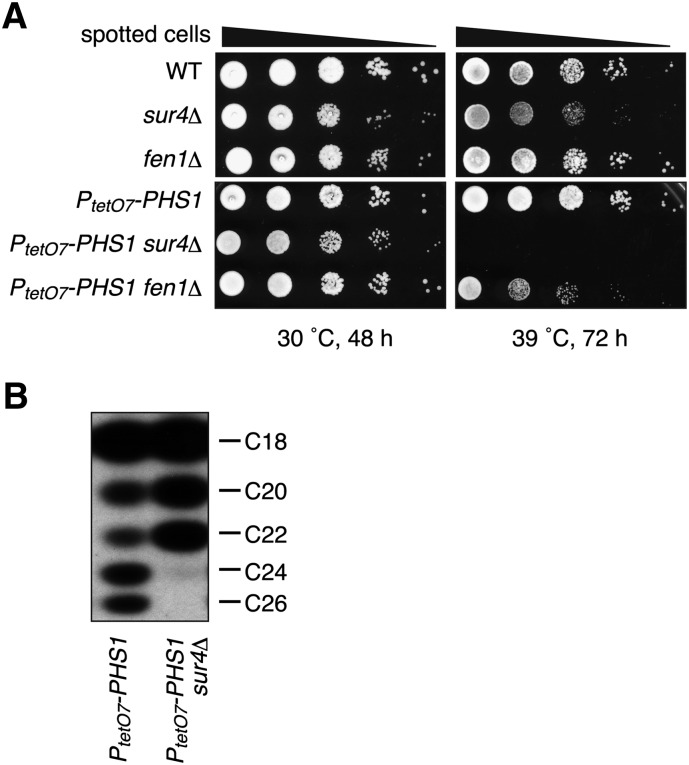
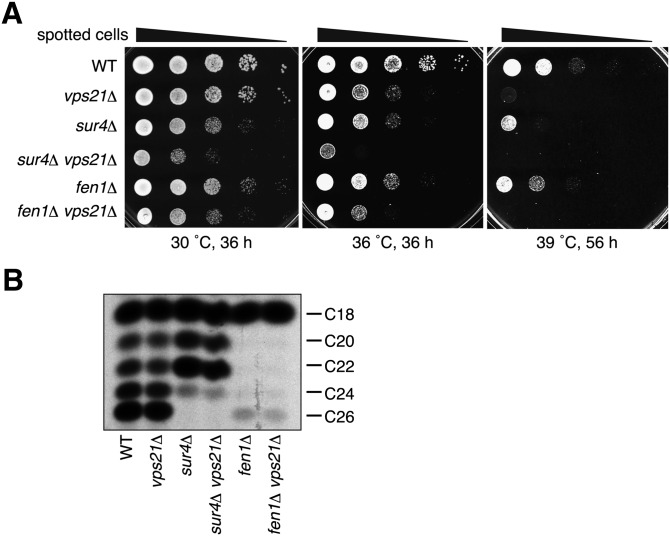
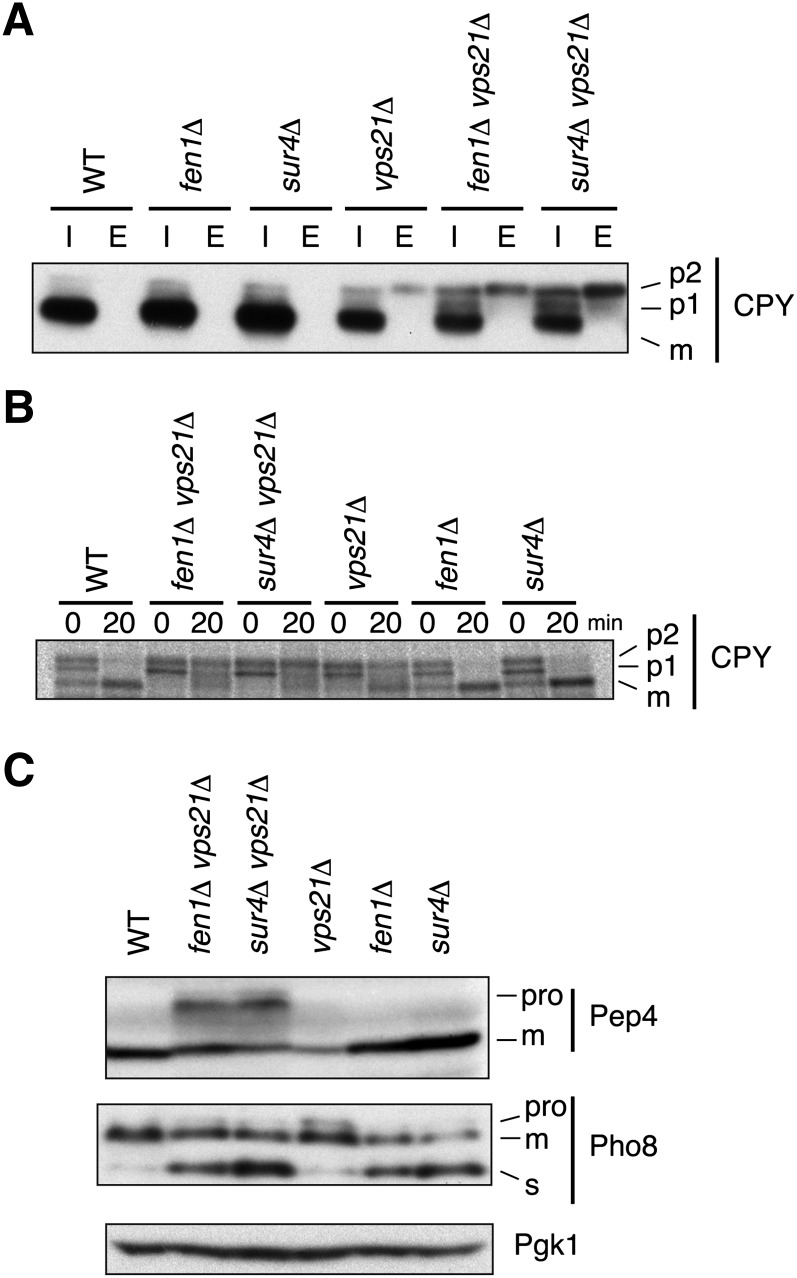
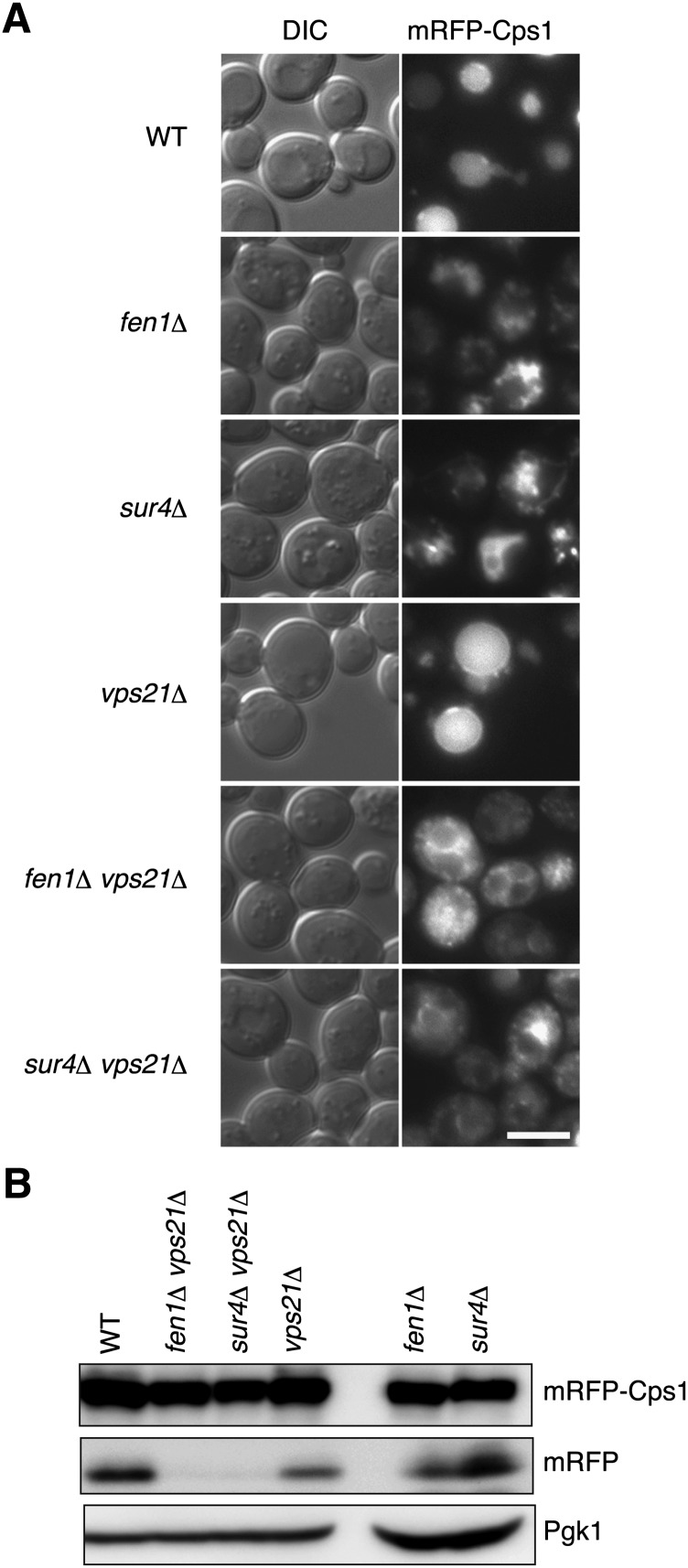
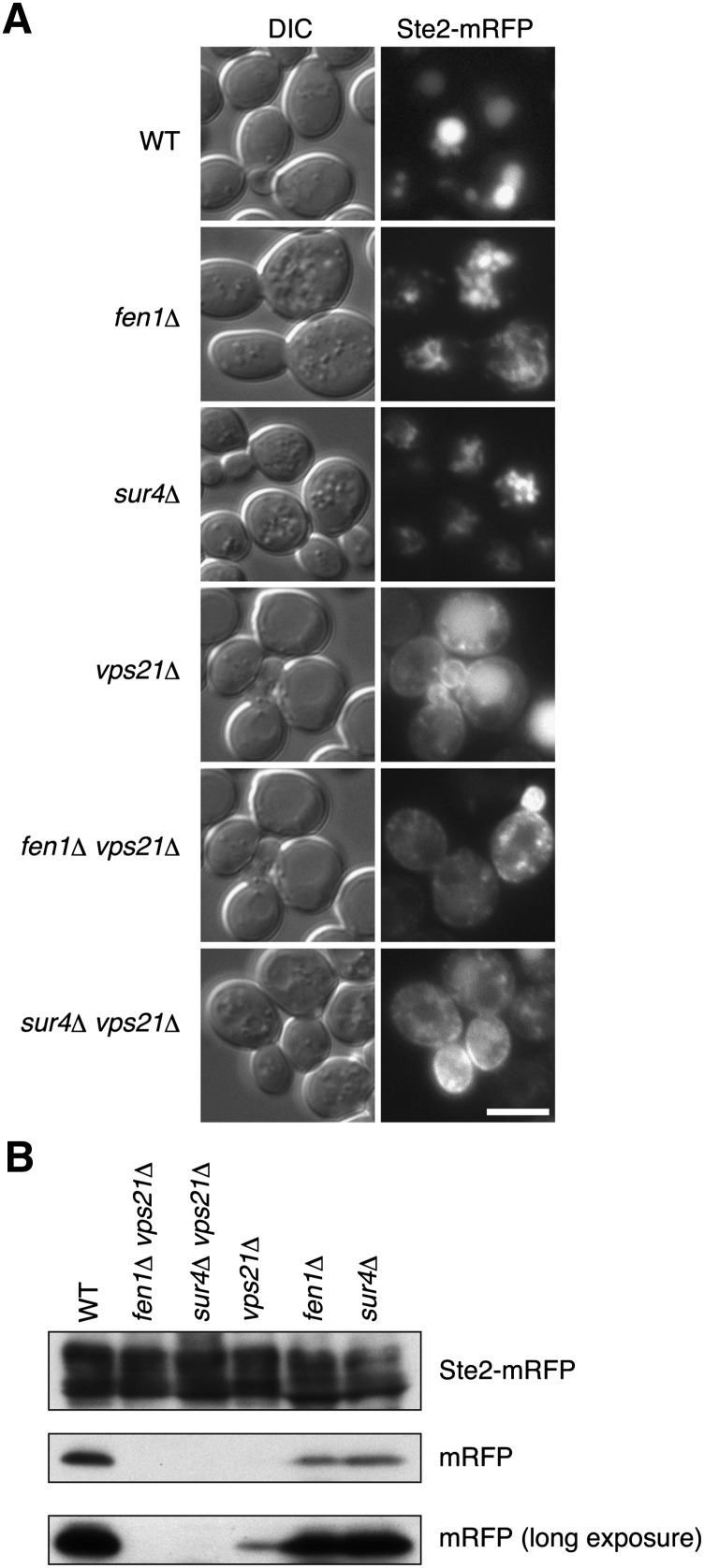
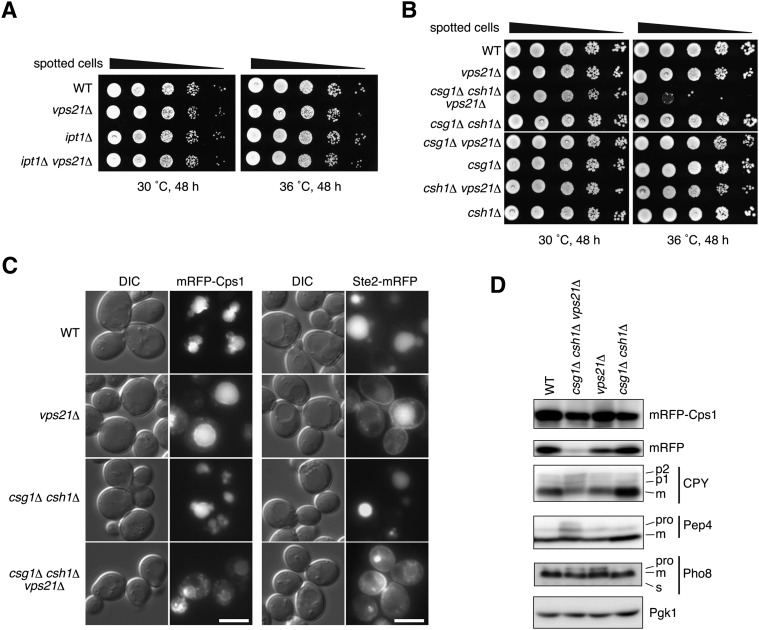
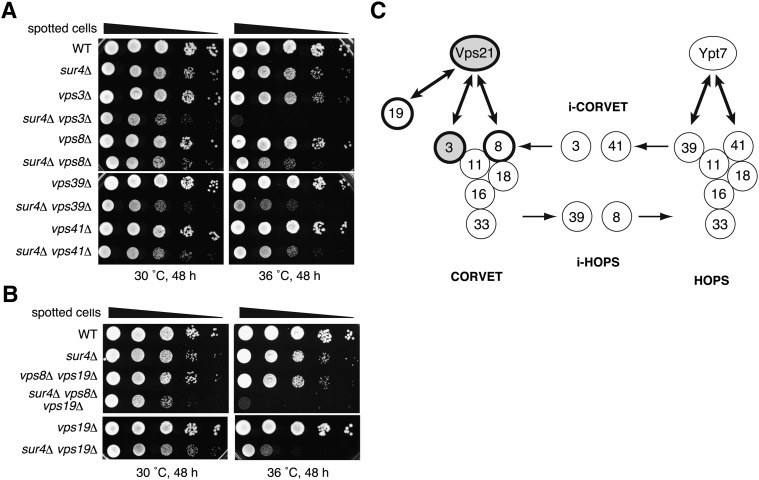
Very long-chain fatty acids (VLCFAs), fatty acids with chain-length greater than 20 carbons, possess a wide range of biological functions. However, their roles at the molecular level remain largely unknown. In the present study, we screened for multicopy suppressors that rescued temperature-sensitive growth of VLCFA-limited yeast cells, and we identified the VPS21 gene, encoding a Rab GTPase, as such a suppressor. When the vps21Δ mutation was introduced into a deletion mutant of the SUR4 gene, which encodes a VLCFA elongase, a synthetic growth defect was observed. Endosome-mediated vesicular trafficking pathways, including endocytosis and the carboxypeptidase Y (CPY) pathway, were severely impaired in sur4Δ vps21Δ double mutants, while the AP-3 pathway that bypasses the endosome was unaffected. In addition, the sur4Δ mutant also exhibited a synthetic growth defect when combined with the deletion of VPS3, which encodes a subunit of the class C core vacuole/endosome tethering (CORVET) complex that tethers transport vesicles to the late endosome/multivesicular body (MVB). These results suggest that, of all the intracellular trafficking pathways, requirement of VLCFAs is especially high in the endosomal pathways.
Figures
Similar articles
-
Functional separation of endosomal fusion factors and the class C core vacuole/endosome tethering (CORVET) complex in endosome biogenesis.J Biol Chem. 2013 Feb 15;288(7):5166-75. doi: 10.1074/jbc.M112.431536. Epub 2012 Dec 21. J Biol Chem. 2013. PMID: 23264632 Free PMC article.
-
Mapping of Vps21 and HOPS binding sites in Vps8 and effect of binding site mutants on endocytic trafficking.Eukaryot Cell. 2010 Apr;9(4):602-10. doi: 10.1128/EC.00286-09. Epub 2010 Feb 19. Eukaryot Cell. 2010. PMID: 20173035 Free PMC article.
-
The N-terminal domains of Vps3 and Vps8 are critical for localization and function of the CORVET tethering complex on endosomes.PLoS One. 2013 Jun 20;8(6):e67307. doi: 10.1371/journal.pone.0067307. Print 2013. PLoS One. 2013. PMID: 23840658 Free PMC article.
-
Tethering complexes in the endocytic pathway: CORVET and HOPS.FEBS J. 2013 Jun;280(12):2743-57. doi: 10.1111/febs.12151. Epub 2013 Feb 21. FEBS J. 2013. PMID: 23351085 Review.
-
Membrane dynamics and fusion at late endosomes and vacuoles--Rab regulation, multisubunit tethering complexes and SNAREs.Eur J Cell Biol. 2011 Sep;90(9):779-85. doi: 10.1016/j.ejcb.2011.04.007. Epub 2011 Jun 16. Eur J Cell Biol. 2011. PMID: 21683469 Review.
Cited by
-
Discovery of broad-spectrum fungicides that block septin-dependent infection processes of pathogenic fungi.Nat Microbiol. 2020 Dec;5(12):1565-1575. doi: 10.1038/s41564-020-00790-y. Epub 2020 Sep 21. Nat Microbiol. 2020. PMID: 32958858
-
The C-terminal Cytosolic Region of Rim21 Senses Alterations in Plasma Membrane Lipid Composition: INSIGHTS INTO SENSING MECHANISMS FOR PLASMA MEMBRANE LIPID ASYMMETRY.J Biol Chem. 2015 Dec 25;290(52):30797-805. doi: 10.1074/jbc.M115.674382. Epub 2015 Nov 2. J Biol Chem. 2015. PMID: 26527678 Free PMC article.
-
Loss of hydroxyl groups from the ceramide moiety can modify the lateral diffusion of membrane proteins in S. cerevisiae.J Lipid Res. 2014 Jul;55(7):1343-56. doi: 10.1194/jlr.M048637. Epub 2014 May 29. J Lipid Res. 2014. PMID: 24875539 Free PMC article.
-
The 3-hydroxyacyl-CoA dehydratases HACD1 and HACD2 exhibit functional redundancy and are active in a wide range of fatty acid elongation pathways.J Biol Chem. 2017 Sep 15;292(37):15538-15551. doi: 10.1074/jbc.M117.803171. Epub 2017 Aug 7. J Biol Chem. 2017. PMID: 28784662 Free PMC article.
-
Dual Fatty Acid Elongase Complex Interactions in Arabidopsis.PLoS One. 2016 Sep 1;11(9):e0160631. doi: 10.1371/journal.pone.0160631. eCollection 2016. PLoS One. 2016. PMID: 27583779 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
