

Selection of binding targets in parasites using phage-display and aptamer libraries in vivo and in vitro
- PMID: 23316203
- PMCID: PMC3540409
- DOI: 10.3389/fimmu.2012.00419
Selection of binding targets in parasites using phage-display and aptamer libraries in vivo and in vitro
Abstract
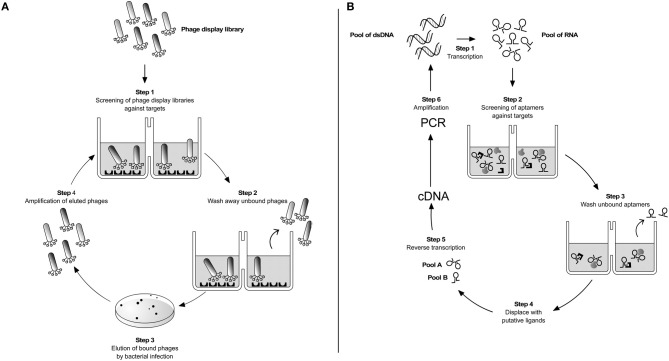
Parasite infections are largely dependent on interactions between pathogen and different host cell populations to guarantee a successful infectious process. This is particularly true for obligatory intracellular parasites as Plasmodium, Toxoplasma, and Leishmania, to name a few. Adhesion to and entry into the cell are essential steps requiring specific parasite and host cell molecules. The large amount of possible involved molecules poses additional difficulties for their identification by the classical biochemical approaches. In this respect, the search for alternative techniques should be pursued. Among them two powerful methodologies can be employed, both relying upon the construction of highly diverse combinatorial libraries of peptides or oligonucleotides that randomly bind with high affinity to targets on the cell surface and are selectively displaced by putative ligands. These are, respectively, the peptide-based phage display and the oligonucleotide-based aptamer techniques. The phage display technique has been extensively employed for the identification of novel ligands in vitro and in vivo in different areas such as cancer, vaccine development, and epitope mapping. Particularly, phage display has been employed in the investigation of pathogen-host interactions. Although this methodology has been used for some parasites with encouraging results, in trypanosomatids its use is, as yet, scanty. RNA and DNA aptamers, developed by the SELEX process (Systematic Evolution of Ligands by Exponential Enrichment), were described over two decades ago and since then contributed to a large number of structured nucleic acids for diagnostic or therapeutic purposes or for the understanding of the cell biology. Similarly to the phage display technique scarce use of the SELEX process has been used in the probing of parasite-host interaction. In this review, an overall survey on the use of both phage display and aptamer technologies in different pathogenic organisms will be discussed. Using these techniques, recent results on the interaction of Trypanosoma cruzi with the host will be highlighted focusing on members of the 85 kDa protein family, a subset of the gp85/TS superfamily.
Keywords: Kinetoplastidae; SELEX; apicomplexa; aptamers; combinatorial methods for diagnosis and therapy; phage display.
Figures
Similar articles
-
The Effects of SELEX Conditions on the Resultant Aptamer Pools in the Selection of Aptamers Binding to Bacterial Cells.J Mol Evol. 2015 Dec;81(5-6):194-209. doi: 10.1007/s00239-015-9711-y. Epub 2015 Nov 4. J Mol Evol. 2015. PMID: 26538121
-
Biological targeting and innovative therapeutic interventions with phage-displayed peptides and structured nucleic acids (aptamers).Curr Opin Biotechnol. 2011 Dec;22(6):832-8. doi: 10.1016/j.copbio.2011.02.012. Epub 2011 Mar 17. Curr Opin Biotechnol. 2011. PMID: 21420292 Review.
-
RNA aptamers: from basic science towards therapy.Handb Exp Pharmacol. 2006;(173):305-26. doi: 10.1007/3-540-27262-3_15. Handb Exp Pharmacol. 2006. PMID: 16594622 Review.
-
In vitro selection of high-affinity nucleic acid ligands to parasite target molecules.Int J Parasitol. 2003 Oct;33(12):1309-17. doi: 10.1016/s0020-7519(03)00197-8. Int J Parasitol. 2003. PMID: 14527514
-
An improved SELEX technique for selection of DNA aptamers binding to M-type 11 of Streptococcus pyogenes.Methods. 2016 Mar 15;97:51-7. doi: 10.1016/j.ymeth.2015.12.005. Epub 2015 Dec 8. Methods. 2016. PMID: 26678795
Cited by
-
Cathepsin L1 mimotopes with adjuvant Quil A induces a Th1/Th2 immune response and confers significant protection against Fasciola hepatica infection in goats.Parasitol Res. 2014 Jan;113(1):243-50. doi: 10.1007/s00436-013-3650-6. Epub 2013 Nov 12. Parasitol Res. 2014. PMID: 24218177
-
Arming Filamentous Bacteriophage, a Nature-Made Nanoparticle, for New Vaccine and Immunotherapeutic Strategies.Pharmaceutics. 2019 Sep 1;11(9):437. doi: 10.3390/pharmaceutics11090437. Pharmaceutics. 2019. PMID: 31480551 Free PMC article. Review.
-
Synthetic, Switchable Enzymes.J Mol Microbiol Biotechnol. 2017;27(2):117-127. doi: 10.1159/000464443. Epub 2017 Apr 28. J Mol Microbiol Biotechnol. 2017. PMID: 28448969 Free PMC article. Review.
-
Phage display screening identifies a novel peptide to suppress ovarian cancer cells in vitro and in vivo in mouse models.BMC Cancer. 2015 Nov 10;15:889. doi: 10.1186/s12885-015-1891-8. BMC Cancer. 2015. PMID: 26555399 Free PMC article.
-
Trypanosoma cruzi-Host Cell Interaction.Front Immunol. 2014 Aug 4;5:339. doi: 10.3389/fimmu.2014.00339. eCollection 2014. Front Immunol. 2014. PMID: 25136341 Free PMC article. No abstract available.
References
-
- Alvarez P., Leguizamon M. S., Buscaglia C. A., Pitcovsky T. A., Campetella O. (2001). Multiple overlapping epitopes in the repetitive unit of the shed acute-phase antigen from Trypanosoma cruzi enhance Its immunogenic properties. Infect. Immun. 69, 7946–7949 10.1128/IAI.69.12.7946-7949.2001 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources