

Functional analysis of the integrator subunit 12 identifies a microdomain that mediates activation of the Drosophila integrator complex
- PMID: 23288851
- PMCID: PMC3576091
- DOI: 10.1074/jbc.M112.425892
Functional analysis of the integrator subunit 12 identifies a microdomain that mediates activation of the Drosophila integrator complex
Abstract
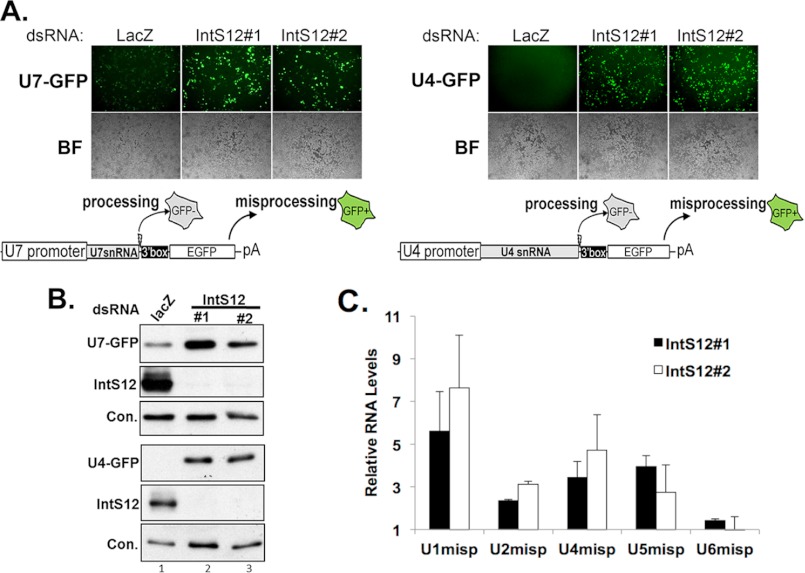
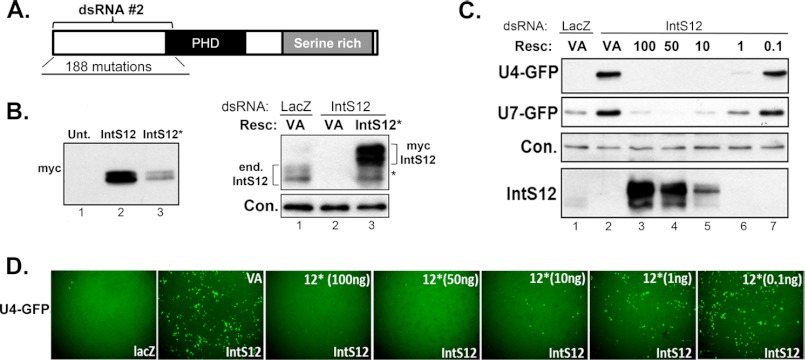
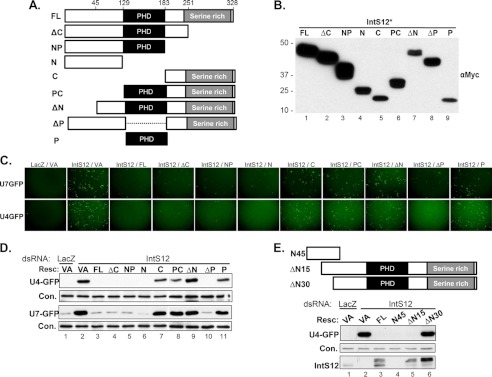
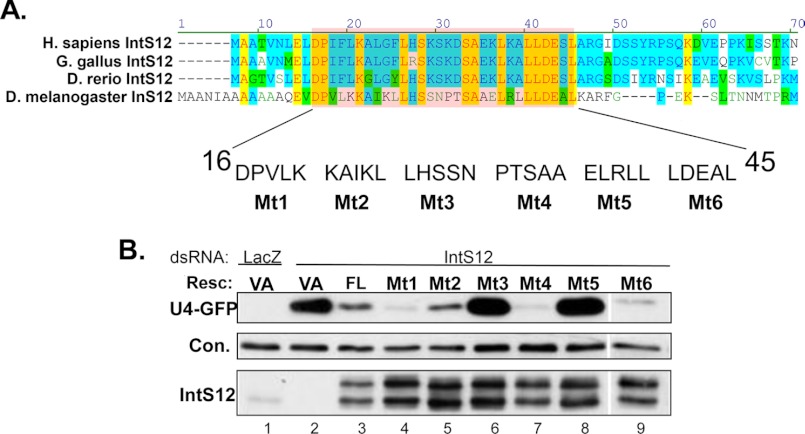
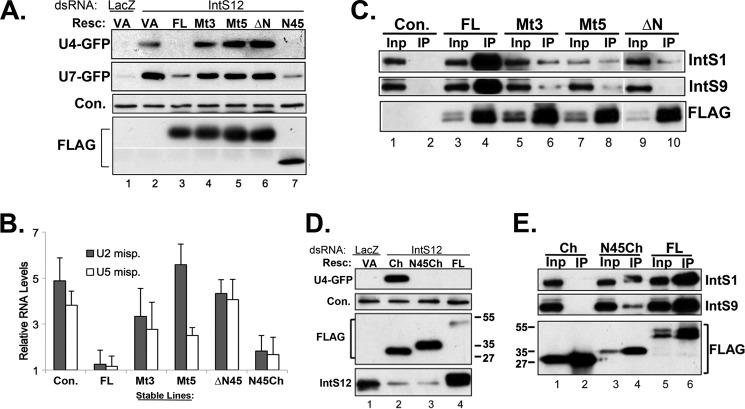
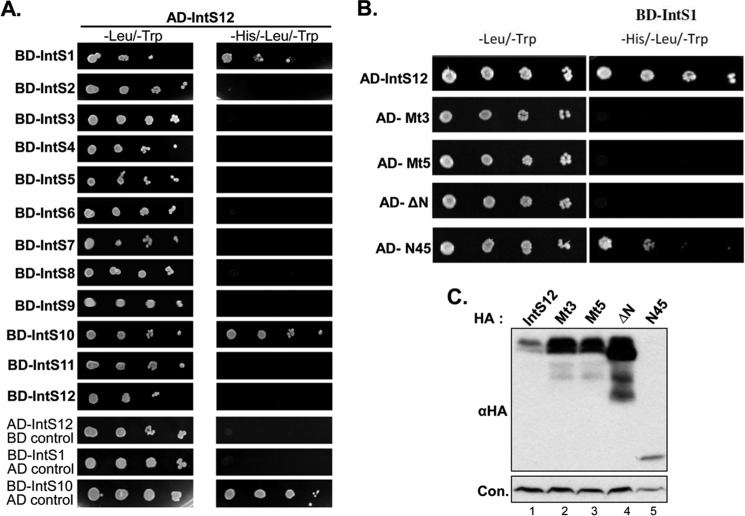
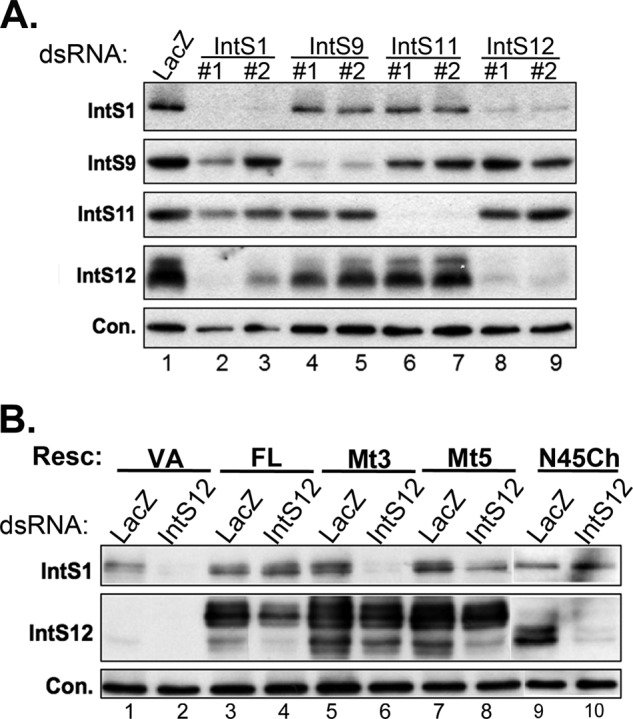
The Drosophila integrator complex consists of 14 subunits that associate with the C terminus of Rpb1 and catalyze the endonucleolytic cleavage of nascent snRNAs near their 3' ends. Although disruption of almost any integrator subunit causes snRNA misprocessing, very little is known about the role of the individual subunits or the network of structural and functional interactions that exist within the complex. Here we developed an RNAi rescue assay in Drosophila S2 cells to identify functional domains within integrator subunit 12 (IntS12) required for snRNA 3' end formation. Surprisingly, the defining feature of the Ints12 protein, a highly conserved and centrally located plant homeodomain finger domain, is not required for reporter snRNA 3' end cleavage. Rather, we find a small, 45-amino acid N-terminal microdomain to be both necessary and nearly sufficient for snRNA biogenesis in cells depleted of endogenous IntS12 protein. This IntS12 microdomain can function autonomously, restoring full integrator processing activity when introduced into a heterologous protein. Moreover, mutations within the microdomain not only disrupt IntS12 function but also abolish binding to other integrator subunits. Finally, the IntS12 microdomain is sufficient to interact and stabilize the putative scaffold integrator subunit, IntS1. Collectively, these results identify an unexpected interaction between the largest and smallest integrator subunits that is essential for the 3' end formation of Drosophila snRNA.
Figures
Similar articles
-
A subset of Drosophila integrator proteins is essential for efficient U7 snRNA and spliceosomal snRNA 3'-end formation.Mol Cell Biol. 2011 Jan;31(2):328-41. doi: 10.1128/MCB.00943-10. Epub 2010 Nov 15. Mol Cell Biol. 2011. PMID: 21078872 Free PMC article.
-
An RNAi screen identifies additional members of the Drosophila Integrator complex and a requirement for cyclin C/Cdk8 in snRNA 3'-end formation.RNA. 2012 Dec;18(12):2148-56. doi: 10.1261/rna.035725.112. Epub 2012 Oct 24. RNA. 2012. PMID: 23097424 Free PMC article.
-
Integrator, a multiprotein mediator of small nuclear RNA processing, associates with the C-terminal repeat of RNA polymerase II.Cell. 2005 Oct 21;123(2):265-76. doi: 10.1016/j.cell.2005.08.019. Cell. 2005. PMID: 16239144
-
snRNA 3' end formation: the dawn of the Integrator complex.Biochem Soc Trans. 2010 Aug;38(4):1082-7. doi: 10.1042/BST0381082. Biochem Soc Trans. 2010. PMID: 20659008 Free PMC article. Review.
-
Regulation of snRNA gene expression by the Drosophila melanogaster small nuclear RNA activating protein complex (DmSNAPc).Crit Rev Biochem Mol Biol. 2011 Feb;46(1):11-26. doi: 10.3109/10409238.2010.518136. Epub 2010 Oct 6. Crit Rev Biochem Mol Biol. 2011. PMID: 20925482 Review.
Cited by
-
Integrator subunit 4 is a 'Symplekin-like' scaffold that associates with INTS9/11 to form the Integrator cleavage module.Nucleic Acids Res. 2018 May 4;46(8):4241-4255. doi: 10.1093/nar/gky100. Nucleic Acids Res. 2018. PMID: 29471365 Free PMC article.
-
Poly(A)-ClickSeq: click-chemistry for next-generation 3΄-end sequencing without RNA enrichment or fragmentation.Nucleic Acids Res. 2017 Jul 7;45(12):e112. doi: 10.1093/nar/gkx286. Nucleic Acids Res. 2017. PMID: 28449108 Free PMC article.
-
IntS6 and the Integrator phosphatase module tune the efficiency of select premature transcription termination events.Mol Cell. 2023 Dec 21;83(24):4445-4460.e7. doi: 10.1016/j.molcel.2023.10.035. Epub 2023 Nov 22. Mol Cell. 2023. PMID: 37995689 Free PMC article.
-
Emerging insights into the function and structure of the Integrator complex.Transcription. 2021 Oct;12(5):251-265. doi: 10.1080/21541264.2022.2047583. Epub 2022 Mar 20. Transcription. 2021. PMID: 35311473 Free PMC article. Review.
-
INTS13 variants causing a recessive developmental ciliopathy disrupt assembly of the Integrator complex.Nat Commun. 2022 Oct 13;13(1):6054. doi: 10.1038/s41467-022-33547-8. Nat Commun. 2022. PMID: 36229431 Free PMC article.
References
-
- Egloff S., O'Reilly D., Murphy S. (2008) Expression of human snRNA genes from beginning to end. Biochem. Soc. Trans. 36, 590–594 - PubMed
-
- Matera A. G., Terns R. M., Terns M. P. (2007) Non-coding RNAs. Lessons from the small nuclear and small nucleolar RNAs. Nat. Rev. Mol. Cell Biol. 8, 209–220 - PubMed
-
- de Vegvar H. E., Lund E., Dahlberg J. E. (1986) 3′ end formation of U1 snRNA precursors is coupled to transcription from snRNA promoters. Cell 47, 259–266 - PubMed
-
- Hernandez N., Weiner A. M. (1986) Formation of the 3′ end of U1 snRNA requires compatible snRNA promoter elements. Cell 47, 249–258 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
