

US28 is a potent activator of phospholipase C during HCMV infection of clinically relevant target cells
- PMID: 23209769
- PMCID: PMC3510093
- DOI: 10.1371/journal.pone.0050524
US28 is a potent activator of phospholipase C during HCMV infection of clinically relevant target cells
Abstract
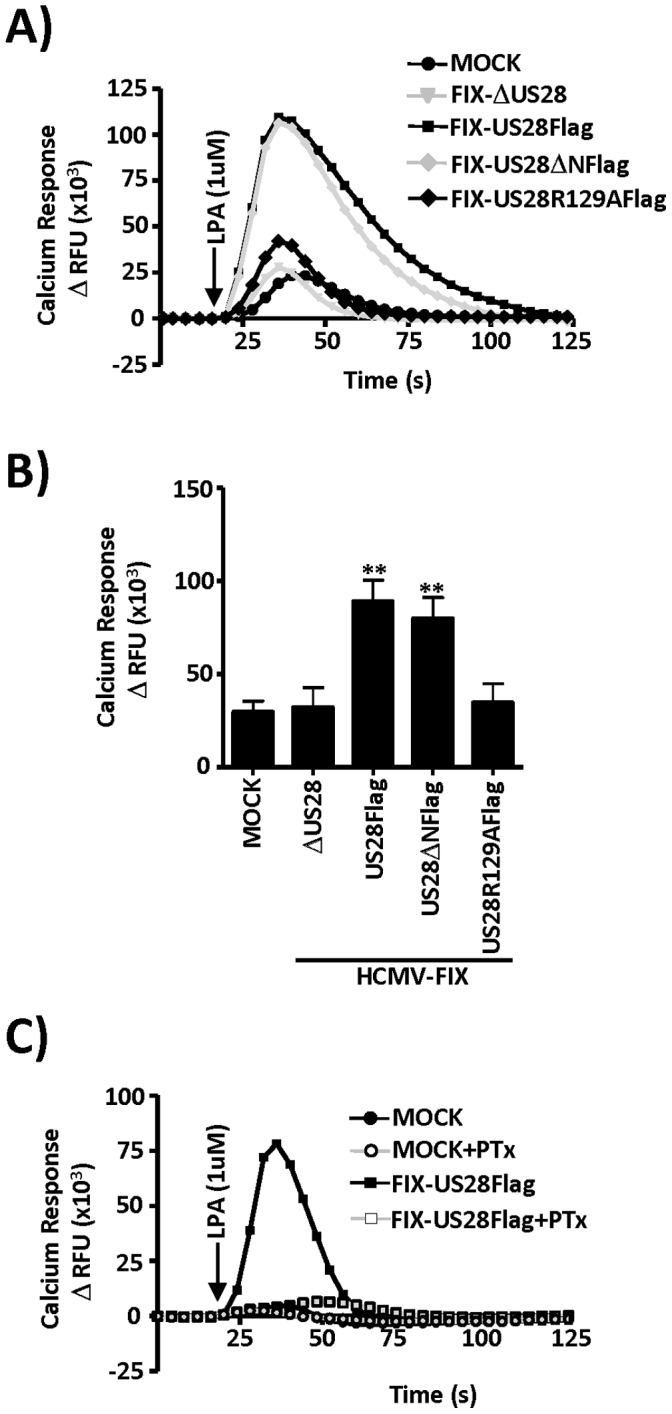
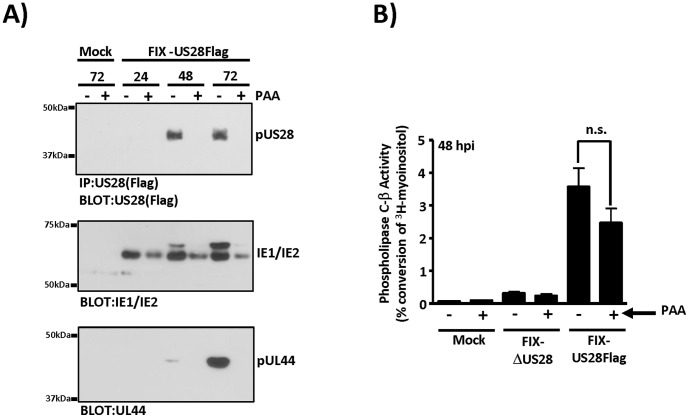
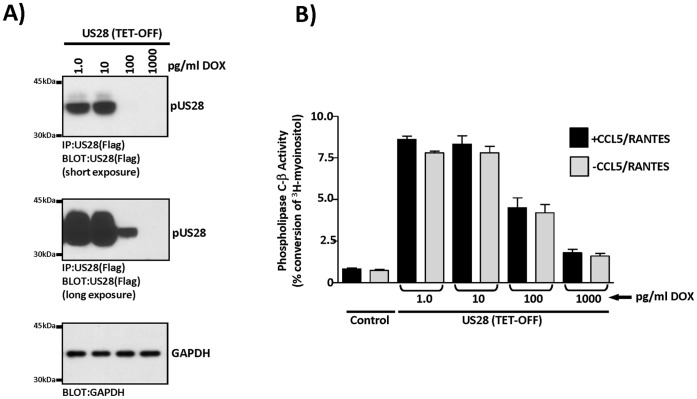
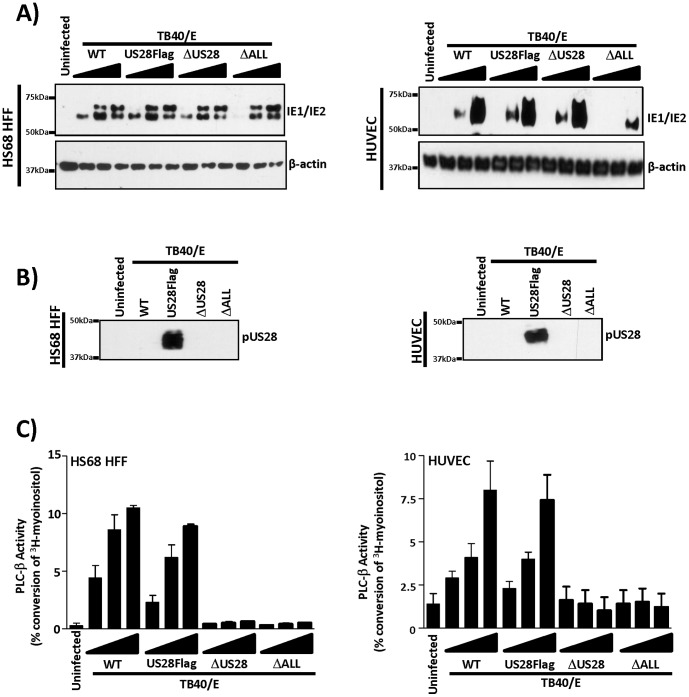
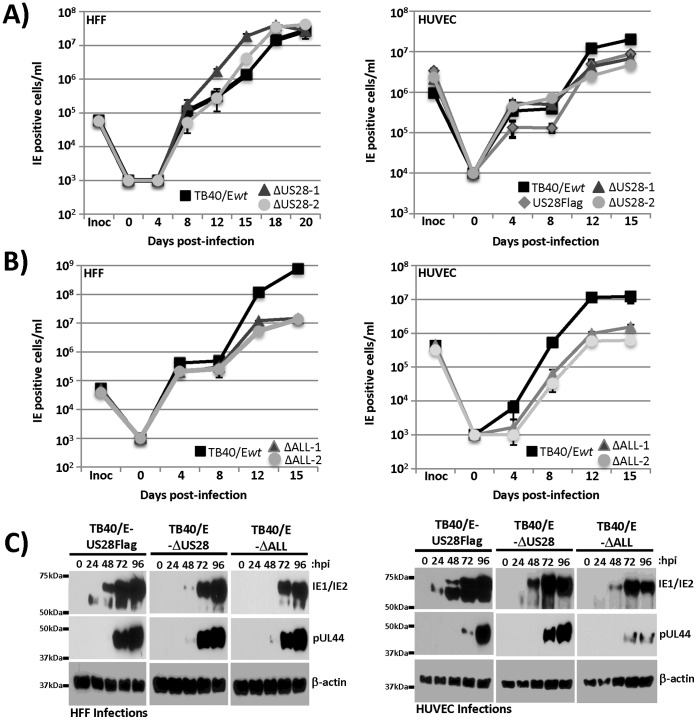
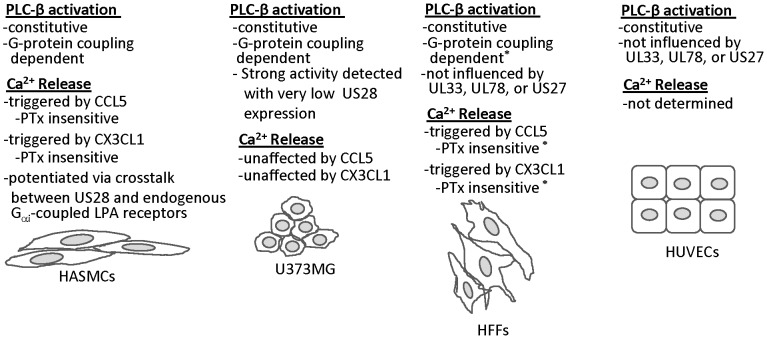
Members of the cytomegalovirus family each encode two or more genes with significant homology to G-protein coupled receptors (GPCRs). In rodent models of pathogenesis, these viral encoded GPCRs play functionally significant roles, as their deletion results in crippled viruses that cannot traffic properly and/or replicate in virally important target cells. Of the four HCMV encoded GPCRs, US28 has garnered the most attention due to the fact that it exhibits both agonist-independent and agonist-dependent signaling activity and has been demonstrated to promote cellular migration and proliferation. Thus, it appears that the CMV GPCRs play important roles in viral replication in vivo as well as promote the development of virus-associated pathology. In the current study we have utilized a series of HCMV/US28 recombinants to investigate the expression profile and signaling activities of US28 in a number of cell types relevant to HCMV infection including smooth muscle cells, endothelial cells and cells derived from glioblastoma multiforme (GBM) tumors. The results indicate that US28 is expressed and exhibits constitutive agonist-independent signaling activity through PLC-β in all cell types tested. Moreover, while CCL5/RANTES and CX3CL1/Fractalkine both promote US28-dependent Ca(++) release in smooth muscle cells, this agonist-dependent effect appears to be cell-specific as we fail to detect US28 driven Ca(++) release in the GBM cells. We have also investigated the effects of US28 on signaling via endogenous GPCRs including those in the LPA receptor family. Our data indicate that US28 can enhance signaling via endogenous LPA receptors. Taken together, our results indicate that US28 induces a variety of signaling events in all cell types tested suggesting that US28 signaling likely plays a significant role during HCMV infection and dissemination in vivo.
Conflict of interest statement
Figures
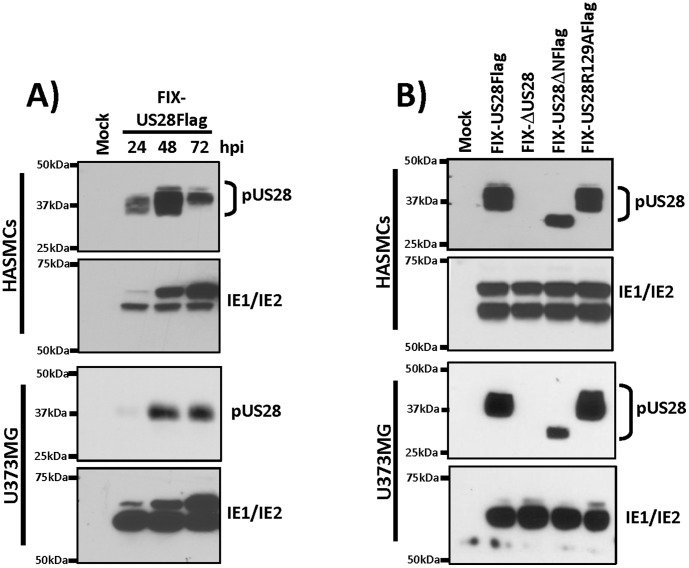
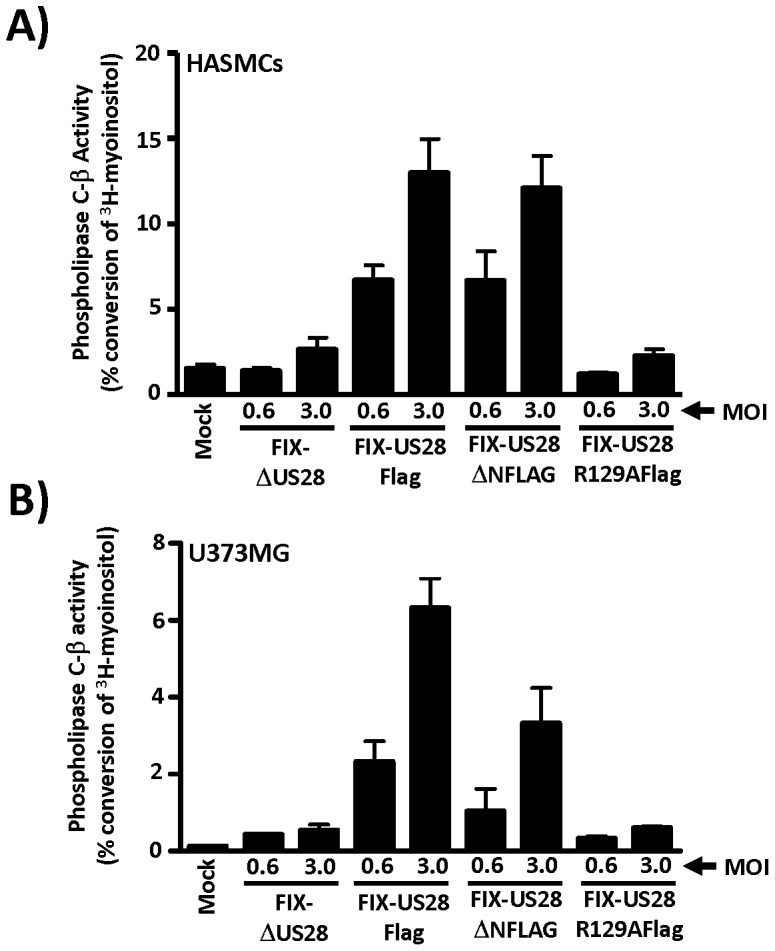
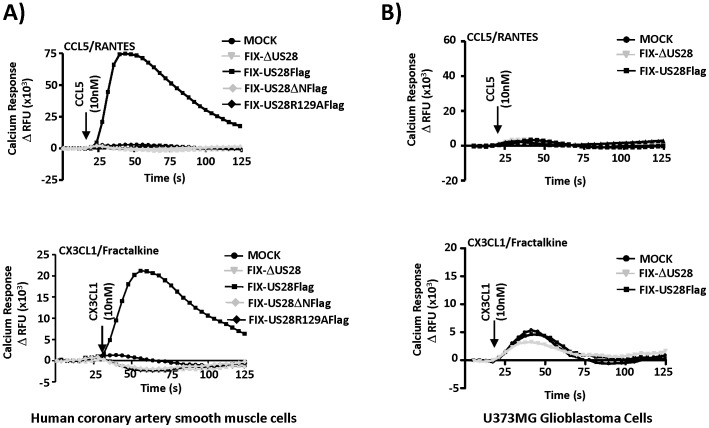
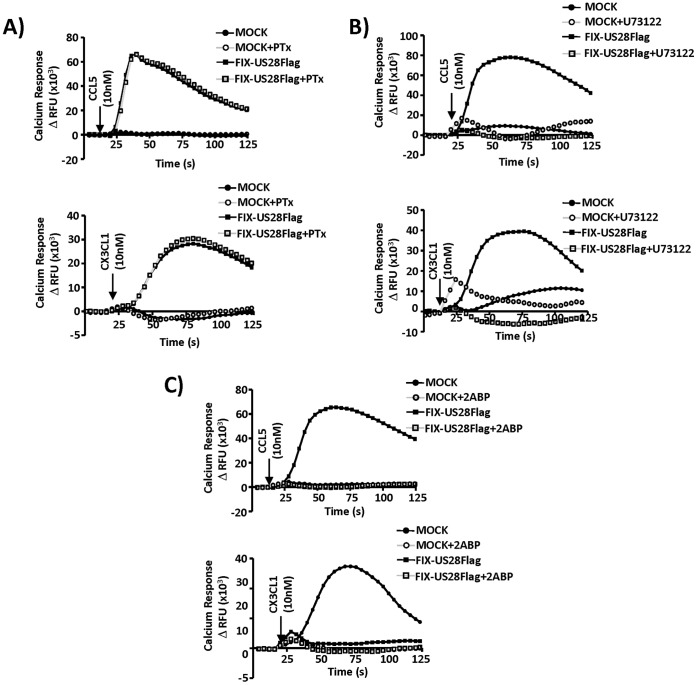
Similar articles
-
The carboxy-terminal tail of human cytomegalovirus (HCMV) US28 regulates both chemokine-independent and chemokine-dependent signaling in HCMV-infected cells.J Virol. 2009 Oct;83(19):10016-27. doi: 10.1128/JVI.00354-09. Epub 2009 Jul 15. J Virol. 2009. PMID: 19605482 Free PMC article.
-
Human cytomegalovirus US28 found in glioblastoma promotes an invasive and angiogenic phenotype.Cancer Res. 2011 Nov 1;71(21):6643-53. doi: 10.1158/0008-5472.CAN-11-0744. Epub 2011 Sep 7. Cancer Res. 2011. PMID: 21900396 Free PMC article.
-
Differential ligand binding to a human cytomegalovirus chemokine receptor determines cell type-specific motility.PLoS Pathog. 2009 Feb;5(2):e1000304. doi: 10.1371/journal.ppat.1000304. Epub 2009 Feb 20. PLoS Pathog. 2009. PMID: 19229316 Free PMC article.
-
Human Cytomegalovirus US28: a functionally selective chemokine binding receptor.Infect Disord Drug Targets. 2009 Nov;9(5):548-56. doi: 10.2174/187152609789105696. Infect Disord Drug Targets. 2009. PMID: 19594424 Free PMC article. Review.
-
US28 actions in HCMV infection: lessons from a versatile hijacker.Rev Med Virol. 2005 Jul-Aug;15(4):269-82. doi: 10.1002/rmv.468. Rev Med Virol. 2005. PMID: 15861487 Review.
Cited by
-
Cytomegalovirus US28 regulates cellular EphA2 to maintain viral latency.Sci Adv. 2022 Oct 28;8(43):eadd1168. doi: 10.1126/sciadv.add1168. Epub 2022 Oct 26. Sci Adv. 2022. PMID: 36288299 Free PMC article.
-
Dysregulation of host cell calcium signaling during viral infections: Emerging paradigm with high clinical relevance.Mol Aspects Med. 2021 Oct;81:101004. doi: 10.1016/j.mam.2021.101004. Epub 2021 Jul 23. Mol Aspects Med. 2021. PMID: 34304899 Free PMC article. Review.
-
Delivery of US28 by incoming HCMV particles rapidly attenuates Akt activity to suppress HCMV lytic replication in monocytes.Sci Signal. 2024 Aug 27;17(851):eadn8727. doi: 10.1126/scisignal.adn8727. Epub 2024 Aug 27. Sci Signal. 2024. PMID: 39190708 Free PMC article.
-
Emerging Role of Compartmentalized G Protein-Coupled Receptor Signaling in the Cardiovascular Field.ACS Pharmacol Transl Sci. 2020 Feb 24;3(2):221-236. doi: 10.1021/acsptsci.0c00006. eCollection 2020 Apr 10. ACS Pharmacol Transl Sci. 2020. PMID: 32296764 Free PMC article. Review.
-
Latency-Associated Expression of Human Cytomegalovirus US28 Attenuates Cell Signaling Pathways To Maintain Latent Infection.mBio. 2017 Dec 5;8(6):e01754-17. doi: 10.1128/mBio.01754-17. mBio. 2017. PMID: 29208743 Free PMC article.
References
-
- Mocarski ES, Courcelle CT (2001) Cytomegaloviruses and their replication. In: Knipe DM, Howley PM, editors. Fields Virology. Philadelpha, PA: Lippinscott Williams & Williams. 2629–2973.
-
- Pass RF (2001) Cytomegaloviruses. In: Knipe DM, Howley PM, editors. Fields Virology. Philadelpha, PA: Lippinscott Williams & Williams. 2675–2706.
-
- Khanna R, Diamond DJ (2006) Human cytomegalovirus vaccine: time to look for alternative options. Trends Mol Med 12: 26–33. - PubMed
-
- Stagno S, Pass RF, Dworsky ME, Henderson RE, Moore EG, et al. (1982) Congenital cytomegalovirus infection: The relative importance of primary and recurrent maternal infection. N Engl J Med 306: 945–949. - PubMed
-
- Boppana SB, Pass RF, Britt WJ, Stagno S, Alford CA (1992) Symptomatic congenital cytomegalovirus infection: neonatal morbidity and mortality. Pediatr Infect Dis J 11: 93–99. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
