

Structural conservation of distinctive N-terminal acetylation-dependent interactions across a family of mammalian NEDD8 ligation enzymes
- PMID: 23201271
- PMCID: PMC3786212
- DOI: 10.1016/j.str.2012.10.013
Structural conservation of distinctive N-terminal acetylation-dependent interactions across a family of mammalian NEDD8 ligation enzymes
Abstract
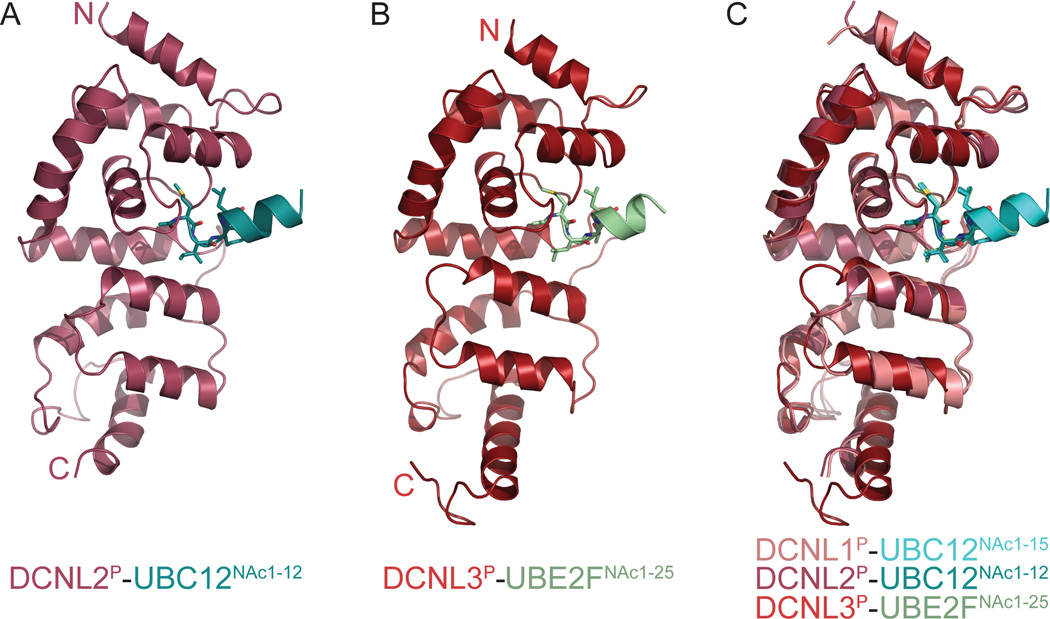
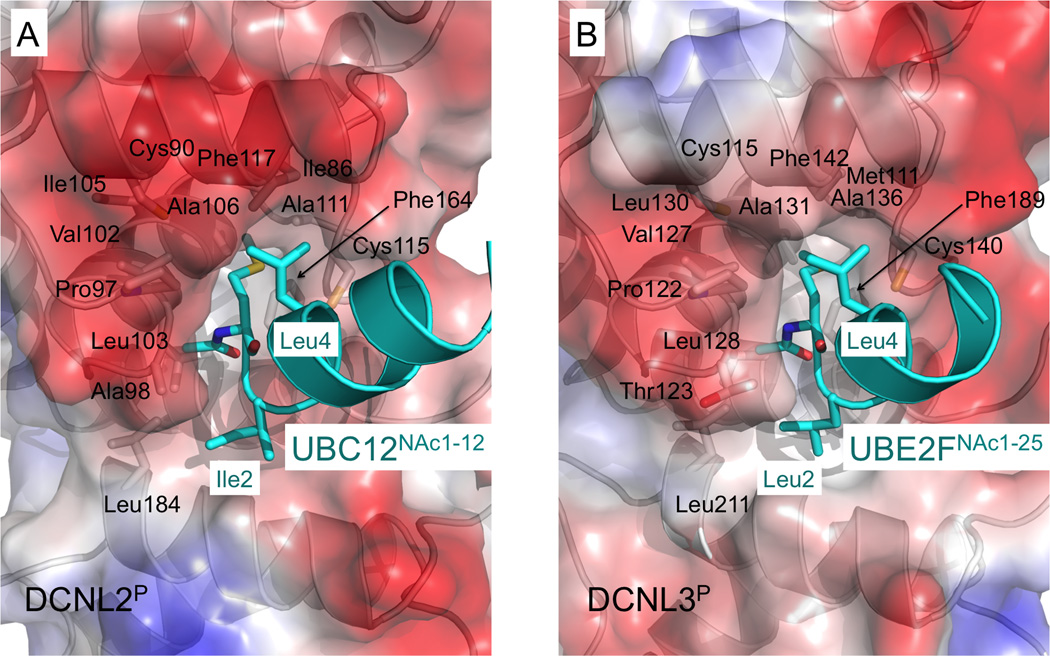
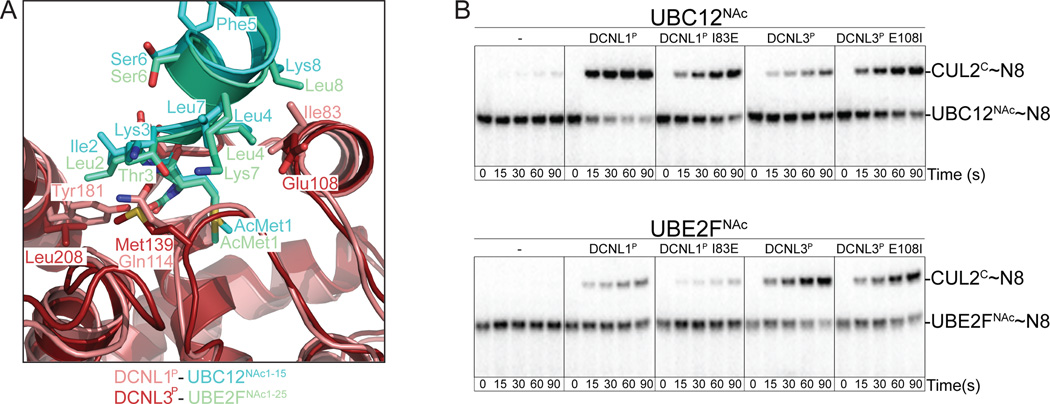
Little is known about molecular recognition of acetylated N termini, despite prevalence of this modification among eukaryotic cytosolic proteins. We report that the family of human DCN-like (DCNL) co-E3s, which promote ligation of the ubiquitin-like protein NEDD8 to cullin targets, recognizes acetylated N termini of the E2 enzymes UBC12 and UBE2F. Systematic biochemical and biophysical analyses reveal 40- and 10-fold variations in affinities among different DCNL-cullin and DCNL-E2 complexes, contributing to varying efficiencies of different NEDD8 ligation cascades. Structures of DCNL2 and DCNL3 complexes with N-terminally acetylated peptides from UBC12 and UBE2F illuminate a common mechanism by which DCNL proteins recognize N-terminally acetylated E2s and how selectivity for interactions dependent on N-acetyl-methionine are established through side chains recognizing distal residues. Distinct preferences of UBC12 and UBE2F peptides for inhibiting different DCNLs, including the oncogenic DCNL1 protein, suggest it may be possible to develop small molecules blocking specific N-acetyl-methionine-dependent protein interactions.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
E2-RING expansion of the NEDD8 cascade confers specificity to cullin modification.Mol Cell. 2009 Feb 27;33(4):483-95. doi: 10.1016/j.molcel.2009.01.011. Mol Cell. 2009. PMID: 19250909 Free PMC article.
-
Biochemical and Structural Consequences of NEDD8 Acetylation.Chembiochem. 2024 Nov 18;25(22):e202400478. doi: 10.1002/cbic.202400478. Epub 2024 Sep 23. Chembiochem. 2024. PMID: 39022855
-
N-terminal acetylation acts as an avidity enhancer within an interconnected multiprotein complex.Science. 2011 Nov 4;334(6056):674-8. doi: 10.1126/science.1209307. Epub 2011 Sep 22. Science. 2011. PMID: 21940857 Free PMC article.
-
Targeting DCN1-UBC12 Protein-Protein Interaction for Regulation of Neddylation Pathway.Adv Exp Med Biol. 2020;1217:349-362. doi: 10.1007/978-981-15-1025-0_20. Adv Exp Med Biol. 2020. PMID: 31898237 Review.
-
Targeting neddylation E2s: a novel therapeutic strategy in cancer.J Hematol Oncol. 2021 Apr 7;14(1):57. doi: 10.1186/s13045-021-01070-w. J Hematol Oncol. 2021. PMID: 33827629 Free PMC article. Review.
Cited by
-
Cross-ancestry analysis identifies genes associated with obesity risk and protection.medRxiv [Preprint]. 2024 Oct 16:2024.10.13.24315422. doi: 10.1101/2024.10.13.24315422. medRxiv. 2024. PMID: 39484254 Free PMC article. Preprint.
-
Ubiquitylation, neddylation and the DNA damage response.Open Biol. 2015 Apr;5(4):150018. doi: 10.1098/rsob.150018. Open Biol. 2015. PMID: 25833379 Free PMC article. Review.
-
Determining the Effects of Neddylation on Cullin-RING Ligase-Dependent Protein Ubiquitination.Curr Protoc. 2022 Mar;2(3):e401. doi: 10.1002/cpz1.401. Curr Protoc. 2022. PMID: 35316580 Free PMC article.
-
N-terminal acetylation and methylation differentially affect the function of MYL9.Biochem J. 2018 Oct 22;475(20):3201-3219. doi: 10.1042/BCJ20180638. Biochem J. 2018. PMID: 30242065 Free PMC article.
-
Overlap of NatA and IAP substrates implicates N-terminal acetylation in protein stabilization.Sci Adv. 2021 Jan 15;7(3):eabc8590. doi: 10.1126/sciadv.abc8590. Print 2021 Jan. Sci Adv. 2021. PMID: 33523899 Free PMC article.
References
-
- Behnia R, Panic B, Whyte JR, Munro S. Targeting of the Arf-like GTPase Arl3p to the Golgi requires N-terminal acetylation and the membrane protein Sys1p. Nat Cell Biol. 2004;6:405–413. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
