

Progenitor and terminal subsets of CD8+ T cells cooperate to contain chronic viral infection
- PMID: 23197535
- PMCID: PMC3653769
- DOI: 10.1126/science.1229620
Progenitor and terminal subsets of CD8+ T cells cooperate to contain chronic viral infection
Abstract
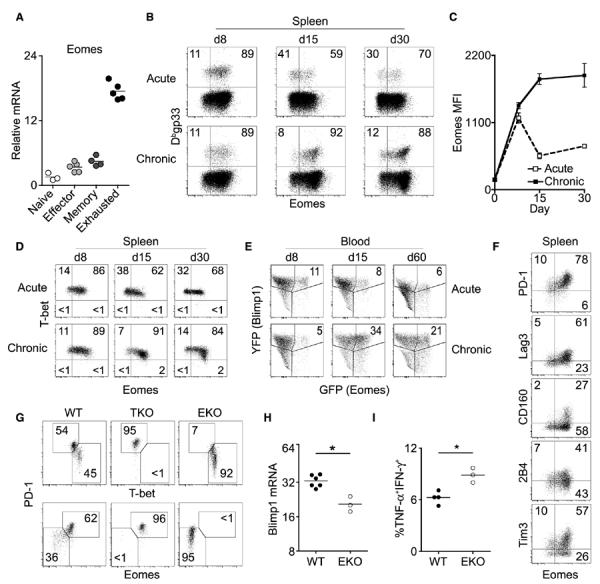
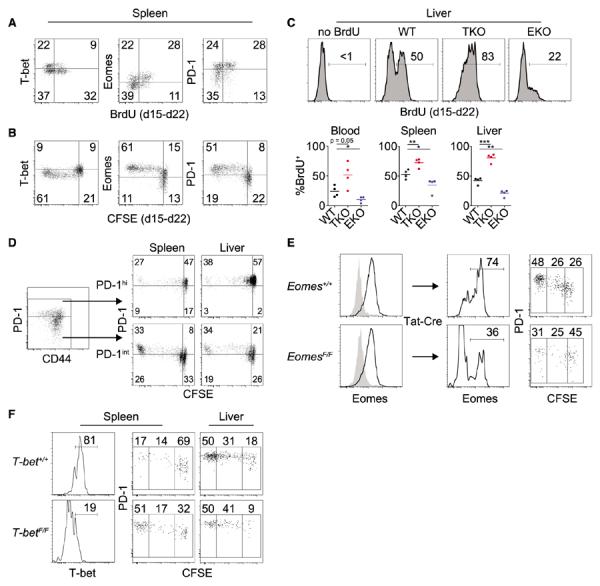
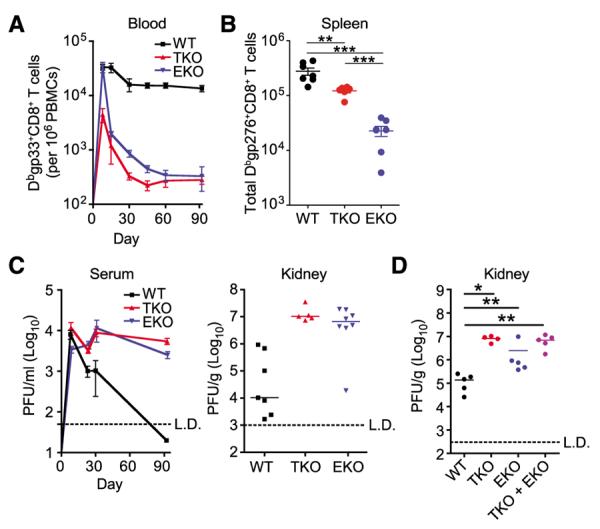
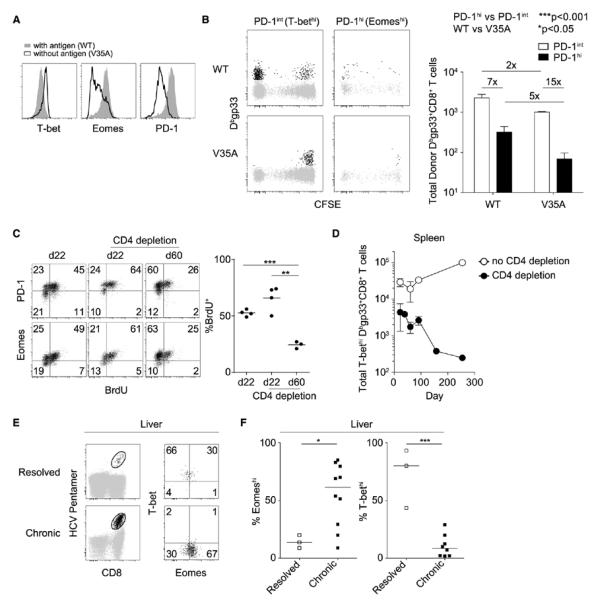
Chronic infections strain the regenerative capacity of antiviral T lymphocyte populations, leading to failure in long-term immunity. The cellular and molecular events controlling this regenerative capacity, however, are unknown. We found that two distinct states of virus-specific CD8(+) T cells exist in chronically infected mice and humans. Differential expression of the T-box transcription factors T-bet and Eomesodermin (Eomes) facilitated the cooperative maintenance of the pool of antiviral CD8(+) T cells during chronic viral infection. T-bet(hi) cells displayed low intrinsic turnover but proliferated in response to persisting antigen, giving rise to Eomes(hi) terminal progeny. Genetic elimination of either subset resulted in failure to control chronic infection, which suggests that an imbalance in differentiation and renewal could underlie the collapse of immunity in humans with chronic infections.
Figures
Comment in
-
T cell responses: Antiviral immunity ticks both T-boxes.Nat Rev Immunol. 2013 Jan;13(1):3. doi: 10.1038/nri3372. Epub 2012 Dec 14. Nat Rev Immunol. 2013. PMID: 23237967 No abstract available.
Similar articles
-
Characterization of eomesodermin and T-bet expression by allostimulated CD8+ T cells of healthy volunteers and kidney transplant patients in relation to graft outcome.Clin Exp Immunol. 2018 Nov;194(2):259-272. doi: 10.1111/cei.13162. Epub 2018 Sep 23. Clin Exp Immunol. 2018. PMID: 30246373 Free PMC article.
-
Endogenous antigen presentation impacts on T-box transcription factor expression and functional maturation of CD8+ T cells.Blood. 2012 Oct 18;120(16):3237-45. doi: 10.1182/blood-2012-03-420182. Epub 2012 Aug 31. Blood. 2012. PMID: 22942184
-
Effect of Cytomegalovirus (CMV) and Ageing on T-Bet and Eomes Expression on T-Cell Subsets.Int J Mol Sci. 2017 Jun 29;18(7):1391. doi: 10.3390/ijms18071391. Int J Mol Sci. 2017. PMID: 28661443 Free PMC article.
-
Infection history determines the differentiation state of human CD8+ T cells.J Virol. 2015 May;89(9):5110-23. doi: 10.1128/JVI.03478-14. Epub 2015 Feb 25. J Virol. 2015. PMID: 25717102 Free PMC article.
-
T-Bet and Eomes Regulate the Balance between the Effector/Central Memory T Cells versus Memory Stem Like T Cells.PLoS One. 2013 Jun 27;8(6):e67401. doi: 10.1371/journal.pone.0067401. Print 2013. PLoS One. 2013. PMID: 23826287 Free PMC article.
Cited by
-
Methylmalonic acid induces metabolic abnormalities and exhaustion in CD8+ T cells to suppress anti-tumor immunity.Oncogene. 2025 Jan;44(2):105-114. doi: 10.1038/s41388-024-03191-1. Epub 2024 Oct 29. Oncogene. 2025. PMID: 39472497
-
Enhancing the Efficacy of CAR-T Cell Therapy: A Comprehensive Exploration of Cellular Strategies and Molecular Dynamics.J Cancer Immunol (Wilmington). 2024;6(1):20-28. doi: 10.33696/cancerimmunol.6.080. J Cancer Immunol (Wilmington). 2024. PMID: 39119270 Free PMC article.
-
LAG-3 sustains TOX expression and regulates the CD94/NKG2-Qa-1b axis to govern exhausted CD8 T cell NK receptor expression and cytotoxicity.Cell. 2024 Aug 8;187(16):4336-4354.e19. doi: 10.1016/j.cell.2024.07.018. Cell. 2024. PMID: 39121847
-
TCR independent suppression of CD8(+) T cell cytokine production mediated by IFNγ in vivo.Virology. 2016 Nov;498:69-81. doi: 10.1016/j.virol.2016.08.003. Epub 2016 Aug 24. Virology. 2016. PMID: 27564543 Free PMC article.
-
The architectural design of CD8+ T cell responses in acute and chronic infection: Parallel structures with divergent fates.J Exp Med. 2021 Apr 5;218(4):e20201730. doi: 10.1084/jem.20201730. J Exp Med. 2021. PMID: 33755719 Free PMC article. Review.
References
-
- Hellerstein M, et al. Nat. Med. 1999;5:83. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- T32 AI007324/AI/NIAID NIH HHS/United States
- U19 AI083022/AI/NIAID NIH HHS/United States
- R56 AI076458/AI/NIAID NIH HHS/United States
- AI082630/AI/NIAID NIH HHS/United States
- AI0663445/AI/NIAID NIH HHS/United States
- T32-AI-07324/AI/NIAID NIH HHS/United States
- R01 AI042370/AI/NIAID NIH HHS/United States
- P30 CA016520/CA/NCI NIH HHS/United States
- R01 AI061699/AI/NIAID NIH HHS/United States
- 059312/WT_/Wellcome Trust/United Kingdom
- AI061699/AI/NIAID NIH HHS/United States
- T32 AI007632/AI/NIAID NIH HHS/United States
- AI083022/AI/NIAID NIH HHS/United States
- AI078897/AI/NIAID NIH HHS/United States
- P30 DK043351/DK/NIDDK NIH HHS/United States
- AI076458/AI/NIAID NIH HHS/United States
- U19 AI082630/AI/NIAID NIH HHS/United States
- P01 AI078897/AI/NIAID NIH HHS/United States
- R01 AI076458/AI/NIAID NIH HHS/United States
- HHSN266200500030C/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
