

Review
doi: 10.1021/cr300360k.
Epub 2012 Nov 14.
Ion channel associated diseases: overview of molecular mechanisms
Affiliations
- PMID: 23151230
- PMCID: PMC3586387
- DOI: 10.1021/cr300360k
Item in Clipboard
Review
Ion channel associated diseases: overview of molecular mechanisms
Chem Rev.
.
No abstract available
Figures
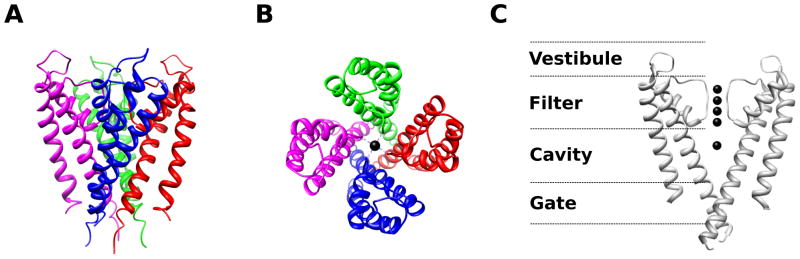
A crystal structure of the KCSA potassium (K+) channel from Streptomyces lividans (PDB 14KC) reveals structural determinants of selectivity and gating. Four KCSA subunits (indicated by different colors) coassemble to form a tetrameric pore structure (A – side view, B – top view). The important structural features are highlighted in panel C – the extracellular vestibule, selectivity filter, pore cavity, and intracellular gate (only two subunits of the tetramer are shown for clarity). Black spheres indicate resolved ions in the filter. The selectivity filter contains a sequence of amino acids (TVGYG) that is highly conserved among K+ channels. The selectivity filter is so narrow that the K+ ions must lose waters of hydration in order to fit. The large energetic penalty associated with desolvation is compensated by interactions with backbone carbonyls of the selectivity filter. Na+ ions do not have the correct geometry to establish stabilizing interactions with the potassium selectivity filter, making the permeability of Na+ through KCSA less likely than K+. In contrast, Rb+, which has a similar geometry as K+, permeates approximately as well as K+ through KCSA,. The presence of multiple ions within the selectivity filter allows for rapid permeation of K+ because the repulsive forces between the ions can dislodge the K+ from the stabilizing interaction sites within the filter. The entry of a new ion into filter pushes the other filter ions along resulting in one ion leaving the filter from the other end. This mode of permeation is analogous to the current in a wire where the rate of current flow is much faster than the drift velocity of individual electrons – even though the entering K+ has only moved part of the way through the filter, one positive charge has been translocated completely across the membrane. The central cavity of the pore is a wide hydrophobic tunnel filled with water molecules that allows for the ion to rejoin with waters of solvation and to continue through the permeation pathway without significant interactions with the protein surface. At the intracellular gate the inner helices of the four subunits form a bundle crossing that creates an impassable constriction of the permeation pathway in the closed state. As revealed by functional studies and the channel structures,–,, local conformational changes around the intracellular gate lead to dilation of the permeation pathway allowing for the passage of ions at the open state. Structural studies show that Na+ and Cl− channels share similar principles of ion selectivity and permeation,–.
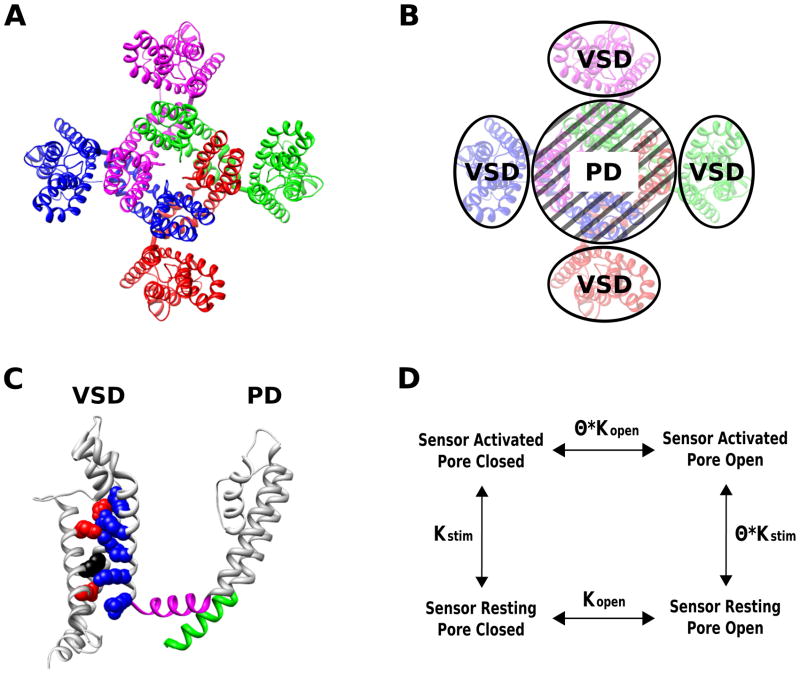
The structure of the voltage-gated, chimeric Kv1.2/2.1 K+ channel reveals the regulation of channel open probability by modular sensor domains. Voltage-gated K+ channels are formed by four subunits (indicated by different colors in A,B) and each of the subunits contains a pore domain and a voltage-sensing domain (VSD) (C). The VSD contains four transmembrane helices (S1–S4), while the pore, like the KCSA pore (Fig. 1), is formed by the coassembly of the pore domains (S5–S6) of all four subunits. The structure clearly reveals the channel’s modular nature consisting of the conserved, central K+ selective pore surrounded by the VSDs (B). VSDs as a structural module have been found in other voltage-gated channels as well as non-ion channel proteins–,–. The VSD contains a series of positively charged residues in the S4 helix, and the membrane potential exerts a force directly upon these charges to displace the S4 segment outward or inward,–. Within the S2 and S3 helices, negatively charged residues are positioned to interact with S4 charges, providing electrostatic forces to stabilize the S4 within the membrane and define the trajectory of the S4 movement,,–. In voltage-gated K+ channels the outward translation of the S4 helix exerts a force on the S6 segment (green) through the S4–S5 linker (magenta) that promotes the dilation of the intracellular gate to open the channel pore–, and S4–S5 linker to gate interactions are found in other voltage-gated channels as well–. Because of these interactions the activation of the VSD and the opening of the pore domain are coupled such that the activation of the sensor increases the probability of the opening of the pore (D). The scheme in panel D represents a general description of the regulation of a channel pore by modular sensor domains. In the scheme, Kstim represents the equilibrium constant for sensor activation which is a function of physiological stimulation, Kopen is the equilibrium constant for the intrinsic pore opening which is independent of the presence of stimulation, θ represents the coupling between the sensor and pore domains whereby sensor activation in response to stimiulation leads to increased pore opening.
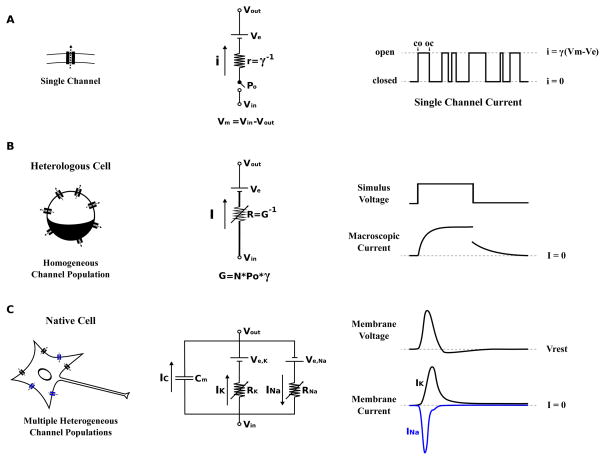
The various scales of electrophysiology. (A) The single ion channel is the molecular determinant of cellular electrophysiology and the therapeutic target for pharmacological approaches to treating channelopathies. At the level of the single channel the current is all or nothing due to the opening and closing of the channel gate(s) (right, CO and OC indicate the first closed to open and open to closed transitions respectively). A circuit model consisting of a battery (Ve), a resistor (r), and a switch (Po) can be used to represent a single ion channel (center). The battery represents the potential generated by asymmetric distribution of permeant ions (the Nernst potential). The selectivity property of the channel determines what ions will permeate and therefore helps to determine the value of Ve. The resistor reflects the ability of the channel to conduct ionic current, resistance is the inverse of conductance (r=γ−1). The switch represents the gating property of the channel through which the conductance can be turned fully on or off (single channel current is all or none). The probability that the switch is open is determined by the probability that the channel gate(s) are open. By convention the membrane potential is defined with respect to the potential inside the membrane (Vm=Vin−Vout). The difference between the membrane potential and the equilibrium potential (Vm−Ve) is called the driving force. This is the equivalent potential that is dropped across the resistor and is therefore linearly related to the magnitude of the current through the resistor by ohms law (Equation 3). (B) Expressing ion channel proteins in heterologous cells, such as Xenopus oocytes and mammalian cell lines (eg. Chinese Hamster Ovary and Human Embryonic Kidney 293 cells), allows for the observation of currents generated by an essentially homogenous population of channels due to relatively low expression of channel proteins in these cells. Macroscopic currents recorded from these cells reflect the summed ensemble of many single channel currents resulting in an apparently smooth response to stimulation. For this reason, the circuit model of a channel population lumps single channel conductance (γ) and the open probability (Po) with the total number of channels (N) into the macroscopic conductance (G). In the circuit model G is represented as a variable conductor as Po changes in response to physiological stimuli (Equation 4). (C) Native cells are yet more complex as they express many different populations of ion channels in the membrane. These channel populations that are coupled through both the membrane potential and physical-chemical environment. Take a simple case of a cell containing only two channel populations; one Na+ channels (Na+ selective), the other K+ channels (K+ permeable), and both are activated by voltage. The opening of the Na+ channels will bias the membrane potential toward the equilibrium potential for Na+ (usually around +50 mV). This depolarization of the membrane potential will open the voltage activated K+ channels that in turn bias the potential back toward the K+ equilibrium potential (usually around −70 mV) meanwhile Na+ channels are closed by inactivation (see main text for the description of inactivation) allowing the K+ channels to dictate the membrane potential. This interplay between the channel populations results in the generation of dynamic electrical signals, known as action potentials, that depend on the properties, shown in Equation 4, for all channels present. Models of these complex systems can be built by connecting the models of individual channel populations (like in B, center) in parallel. In all these circuit models (A–C) the ability of the membrane to separate charged ions is represented as a capacitor in parallel (as shown in C, center).
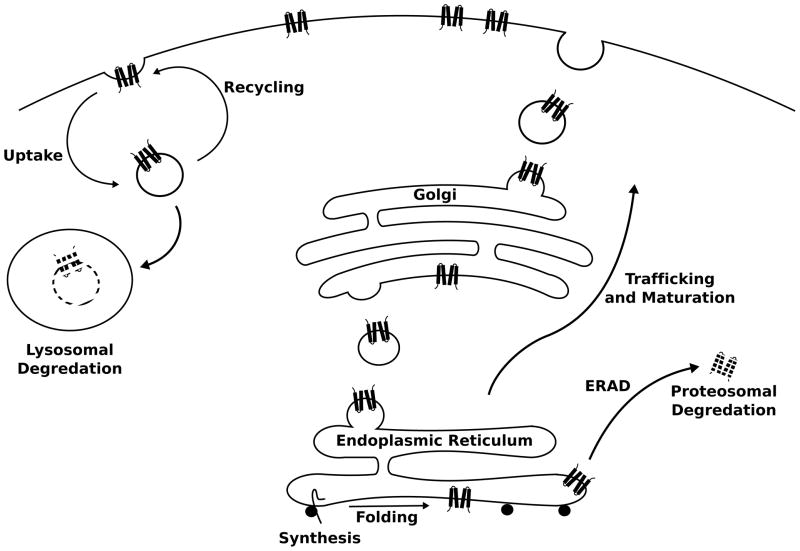
Metabolism of ion channel proteins. Ion channels are synthesized into and across the membrane of the rough endoplasmic reticulum (ER). Channels that achieve a proper fold go on to traffick through the ER and Golgi comparment while undergoing post-translational modifications en route to the target membrane. Improperly folded channels are recognized by the ER quality control machinery and targeted for removal from the ER membrane and proteasomal degredation within the cytosol. After a period of residency within the target membrane, mature channels are uptaken from the membrane into vesicles that can be later returned to the membrane (recycling) or targeted to the lysosome for degredation.
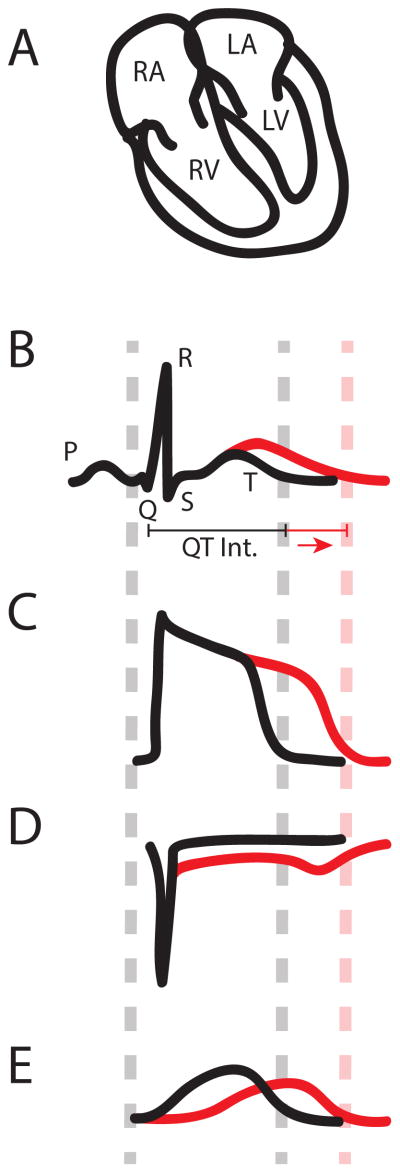
QT interval and action potential (AP) prolongation caused by genetic mutations. The ECG measures the electrical activity of the heart reflected on the body surface, and the waveform of ECG correlates with electrical depolarization and repolarization of the cardiac muscle in various chambers (A, B). The first upward deflection in panel B (P wave) corresponds to activation of the upper heart chambers, the left and right atria (LA and RA, respectively), which collect blood that is returning from the body (RA) and the lungs (LA). The prominent spike formed by the Q, R, and S points is linked to excitation of the massive lower chambers, the left and right ventricles (LV and RV, respectively). The magnitude of the QRS complex is a consequence of the larger ventricular muscle mass, which is needed to generate the force that pushes blood through the body (LV) and the lungs (RV). Finally, the T wave occurs when the ventricles return to an electrical resting state. Consequently, we would expect that a prolonged QT interval is caused by an increase in the time that the ventricles remain in an electrically excited state. The individual cells in the myocardium, myocytes, each generate an action potential (AP) (C) that is responsible for excitation. Therefore, the QT interval (B) corresponds to the duration of the ventricular AP (C), implying that any change in AP duration (APD) will affect the QT interval. APD is determined by a delicate balance of inward and outward ionic currents. The morphology of the AP (C) is the consequence of positively charged Na+, Ca2+ and K+ ions entering and exiting the myocyte. For example, as the membrane potential (Vm) rises from its resting state, caused by the excitation of a neighboring myocytes, Na+ channels open and positively charged Na+ ions enter the cell (D). This inward sodium current (INa) causes Vm to rise quickly. Once Vm is elevated by INa, voltage-gated L-type Ca2+ channels open and bring in a sustained inward Ca2+ flux, ICa,L
(not shown). It is this influx of Ca2+ that signals contraction of the myocyte. The sustained ICa,L supports the AP plateau, which continues until the K+ channels open to generate repolarizing outward current (E). Throughout the AP a small inward Na+ current persists and is enhanced towared the end of the AP when channels begin to recover from inactivation, but are not deactivated yet (window current). In ventricular myocytes there are two major repolarizing K+ currents, one rapid component (IKr) and one slow (IKs). The IKr α-subunit, Kv11.1, is encoded by the KCNH2 (aka HERG) gene and the IKs α-subunit, Kv7.1, is encoded by the KCNQ1 (aka KvLQT1) gene–. Once the outward K+ flux overwhelms the inward Ca2+ flux, the myocyte returns to its resting state. APD is therefore determined by the balance of inward and outward ion fluxes. Increasing inward currents (D) or reducing outward currents (E) will prolong APD (C) and therefore the QT interval (B).
Similar articles
-
SnapShot: Channel Gating Mechanisms.Cell. 2017 Jul 27;170(3):594-594.e1. doi: 10.1016/j.cell.2017.07.019. Cell. 2017. PMID: 28753432
-
From molecule to malady.Nature. 2006 Mar 23;440(7083):440-7. doi: 10.1038/nature04707. Nature. 2006. PMID: 16554803 Review.
-
Structure and function of voltage-gated ion channels.Annu Rev Biochem. 1995;64:493-531. doi: 10.1146/annurev.bi.64.070195.002425. Annu Rev Biochem. 1995. PMID: 7574491 Review.
-
Ion channel-directed therapies in autonomic neurons: a view from the bench.Clin Auton Res. 2002 Apr;12(2):59-65. doi: 10.1007/s102860200021. Clin Auton Res. 2002. PMID: 12102451 Review.
-
Engineering light-gated ion channels.Biochemistry. 2006 Dec 26;45(51):15129-41. doi: 10.1021/bi0618058. Epub 2006 Dec 2. Biochemistry. 2006. PMID: 17176035 Review.
Cited by
-
Electronic Polarization Leads to a Drier Dewetted State for Hydrophobic Gating in the Big Potassium Channel.J Phys Chem Lett. 2024 Jul 25;15(29):7436-7441. doi: 10.1021/acs.jpclett.4c01359. Epub 2024 Jul 15. J Phys Chem Lett. 2024. PMID: 39008088
-
Inner pore hydration free energy controls the activation of big potassium channels.Biophys J. 2023 Apr 4;122(7):1158-1167. doi: 10.1016/j.bpj.2023.02.005. Epub 2023 Feb 10. Biophys J. 2023. PMID: 36774534 Free PMC article.
-
Recent Advances in Computer-Aided Structure-Based Drug Design on Ion Channels.Int J Mol Sci. 2023 May 25;24(11):9226. doi: 10.3390/ijms24119226. Int J Mol Sci. 2023. PMID: 37298178 Free PMC article. Review.
-
A computational model of organism development and carcinogenesis resulting from cells' bioelectric properties and communication.Sci Rep. 2022 Jun 2;12(1):9206. doi: 10.1038/s41598-022-13281-3. Sci Rep. 2022. PMID: 35654933 Free PMC article.
-
Waixenicin A, a marine-derived TRPM7 inhibitor: a promising CNS drug lead.Acta Pharmacol Sin. 2020 Dec;41(12):1519-1524. doi: 10.1038/s41401-020-00512-4. Epub 2020 Sep 29. Acta Pharmacol Sin. 2020. PMID: 32994545 Free PMC article. Review.
References
-
- Armstrong CM, Hille B. Neuron. 1998;20:371. - PubMed
-
- Yang J, Ellinor PT, Sather WA, Zhang JF, Tsien RW. Nature. 1993;366:158. - PubMed
-
- Doyle DA, Morais Cabral J, Pfuetzner RA, Kuo A, Gulbis JM, Cohen SL, Chait BT, MacKinnon R. Science. 1998;280:69. - PubMed
-
- Zhou Y, Morais-Cabral JH, Kaufman A, MacKinnon R. Nature. 2001;414:43. - PubMed
-
- MacKinnon R. Angewandte Chemie. 2004;43:4265. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
