

Caenorhabditis elegans HSF-1 is an essential nuclear protein that forms stress granule-like structures following heat shock
- PMID: 23107491
- PMCID: PMC3552056
- DOI: 10.1111/acel.12024
Caenorhabditis elegans HSF-1 is an essential nuclear protein that forms stress granule-like structures following heat shock
Abstract
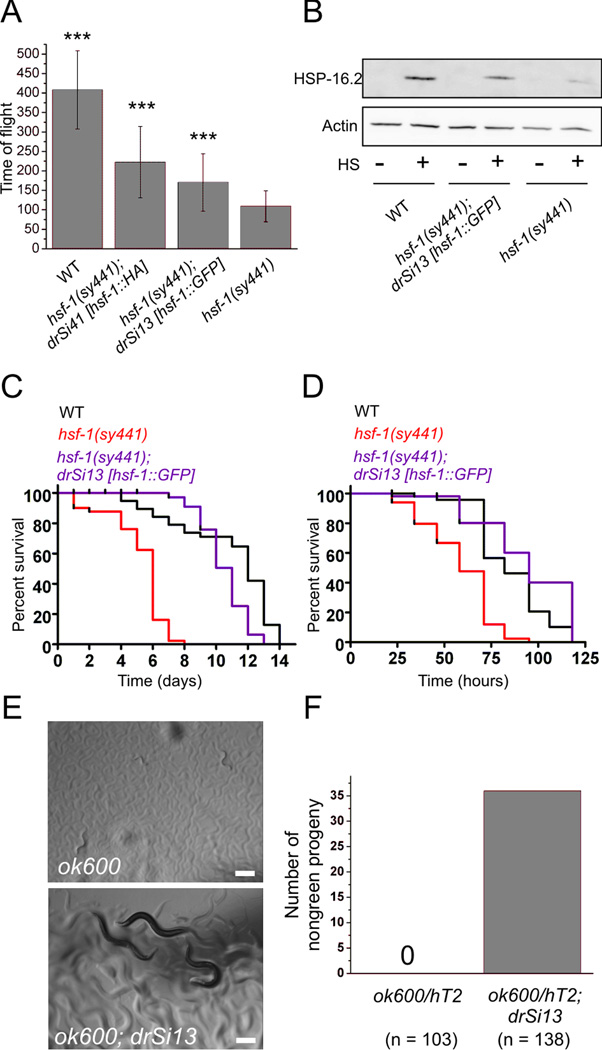
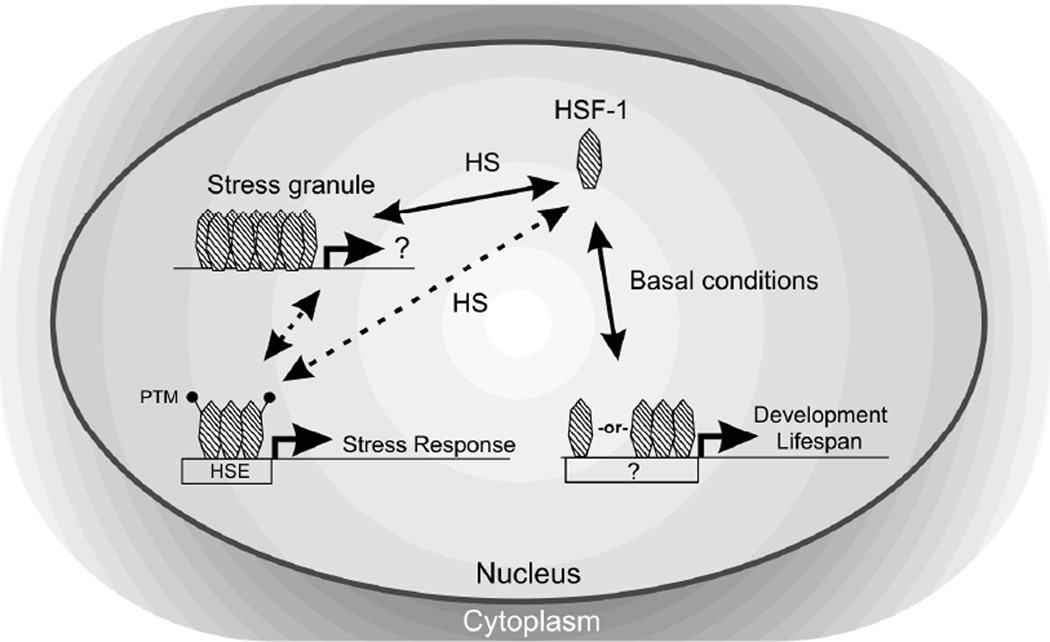
The heat shock transcription factor (HSF) is a conserved regulator of heat shock-inducible gene expression. Organismal roles for HSF in physiological processes such as development, aging, and immunity have been defined largely through studies of the single Caenorhabditis elegans HSF homolog, hsf-1. However, the molecular and cell biological properties of hsf-1 in C. elegans are incompletely understood. We generated animals expressing physiological levels of an HSF-1::GFP fusion protein and examined its function, localization, and regulation in vivo. HSF-1::GFP was functional, as measured by its ability to rescue phenotypes associated with two hsf-1 mutant alleles. Rescue of hsf-1 development phenotypes was abolished in a DNA-binding-deficient mutant, demonstrating that the transcriptional targets of hsf-1 are critical to its function even in the absence of stress. Under nonstress conditions, HSF-1::GFP was found primarily in the nucleus. Following heat shock, HSF-1::GFP rapidly and reversibly redistributed into dynamic, subnuclear structures that share many properties with human nuclear stress granules, including colocalization with markers of active transcription. Rapid formation of HSF-1 stress granules required HSF-1 DNA-binding activity, and the threshold for stress granule formation was altered by growth temperature. HSF-1 stress granule formation was not induced by inhibition of IGF signaling, a pathway previously suggested to function upstream of hsf-1. Our findings suggest that development, stress, and aging pathways may regulate HSF-1 function in distinct ways, and that HSF-1 nuclear stress granule formation is an evolutionarily conserved aspect of HSF-1 regulation in vivo.
© 2012 The Authors Aging Cell © 2012 Blackwell Publishing Ltd/Anatomical Society of Great Britain and Ireland.
Figures




Similar articles
-
The genome-wide role of HSF-1 in the regulation of gene expression in Caenorhabditis elegans.BMC Genomics. 2016 Aug 5;17:559. doi: 10.1186/s12864-016-2837-5. BMC Genomics. 2016. PMID: 27496166 Free PMC article.
-
HSB-1 Inhibition and HSF-1 Overexpression Trigger Overlapping Transcriptional Changes To Promote Longevity in Caenorhabditis elegans.G3 (Bethesda). 2019 May 7;9(5):1679-1692. doi: 10.1534/g3.119.400044. G3 (Bethesda). 2019. PMID: 30894454 Free PMC article.
-
The homeodomain-interacting protein kinase HPK-1 preserves protein homeostasis and longevity through master regulatory control of the HSF-1 chaperone network and TORC1-restricted autophagy in Caenorhabditis elegans.PLoS Genet. 2017 Oct 16;13(10):e1007038. doi: 10.1371/journal.pgen.1007038. eCollection 2017 Oct. PLoS Genet. 2017. PMID: 29036198 Free PMC article.
-
The Thermal Stress Coping Network of the Nematode Caenorhabditis elegans.Int J Mol Sci. 2022 Nov 28;23(23):14907. doi: 10.3390/ijms232314907. Int J Mol Sci. 2022. PMID: 36499234 Free PMC article. Review.
-
Heat shock and genetic activation of HSF-1 enhance immunity to bacteria.Cell Cycle. 2006 Nov 1;5(21):2443-6. doi: 10.4161/cc.5.21.3434. Epub 2006 Sep 25. Cell Cycle. 2006. PMID: 17106259 Review.
Cited by
-
Aging causes decreased resistance to multiple stresses and a failure to activate specific stress response pathways.Aging (Albany NY). 2016 Apr;8(4):777-95. doi: 10.18632/aging.100939. Aging (Albany NY). 2016. PMID: 27053445 Free PMC article.
-
Phycoerythrin extends life span and health span of Caenorhabditis elegans.Age (Dordr). 2014;36(5):9717. doi: 10.1007/s11357-014-9717-1. Epub 2014 Oct 11. Age (Dordr). 2014. PMID: 25304463 Free PMC article.
-
Environmental Canalization of Life Span and Gene Expression in Caenorhabditis elegans.J Gerontol A Biol Sci Med Sci. 2017 Aug 1;72(8):1033-1037. doi: 10.1093/gerona/glx017. J Gerontol A Biol Sci Med Sci. 2017. PMID: 28369388 Free PMC article.
-
Ethanol Stimulates Locomotion via a Gαs-Signaling Pathway in IL2 Neurons in Caenorhabditis elegans.Genetics. 2017 Nov;207(3):1023-1039. doi: 10.1534/genetics.117.300119. Epub 2017 Sep 26. Genetics. 2017. PMID: 28951527 Free PMC article.
-
Three Rules Explain Transgenerational Small RNA Inheritance in C. elegans.Cell. 2020 Sep 3;182(5):1186-1197.e12. doi: 10.1016/j.cell.2020.07.022. Epub 2020 Aug 24. Cell. 2020. PMID: 32841602 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
