

Seven functional classes of Barth syndrome mutation
- PMID: 23100323
- PMCID: PMC3606006
- DOI: 10.1093/hmg/dds447
Seven functional classes of Barth syndrome mutation
Abstract
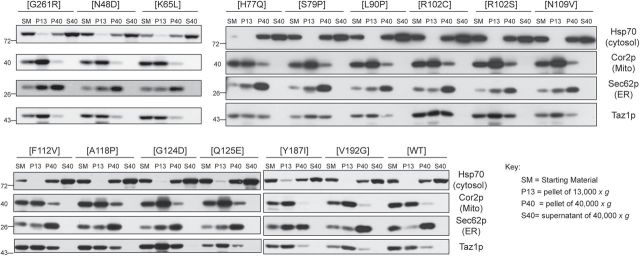
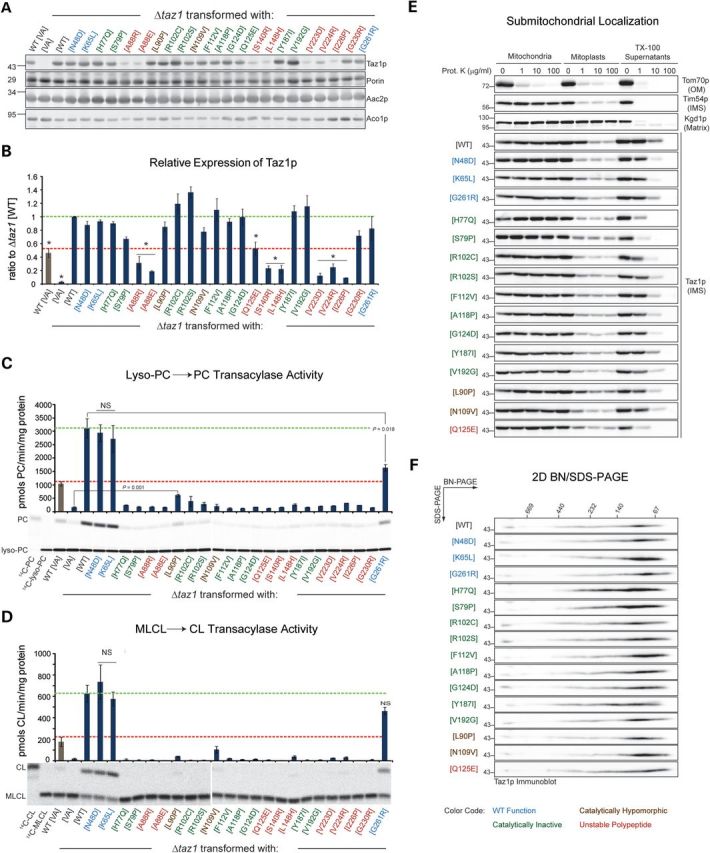
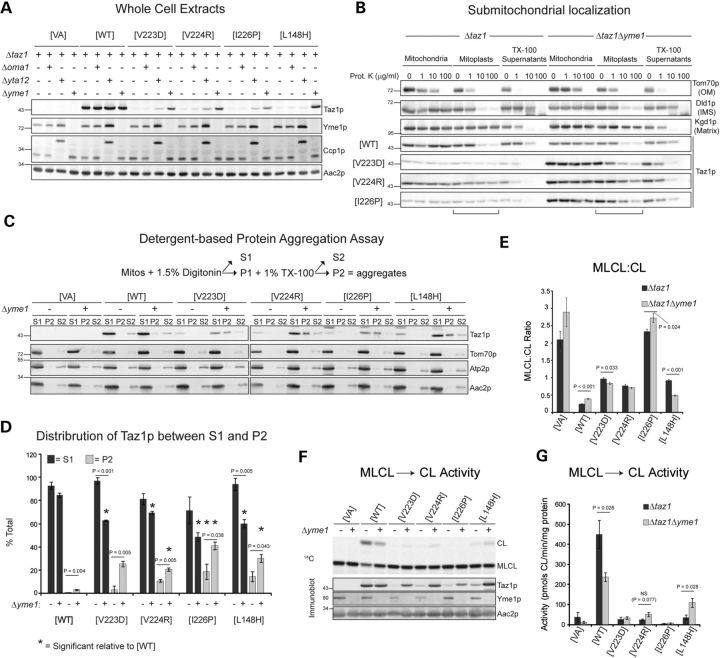
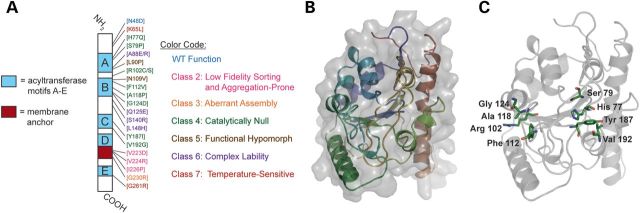
Patients with Barth syndrome (BTHS), a rare X-linked disease, suffer from skeletal and cardiomyopathy and bouts of cyclic neutropenia. The causative gene encodes tafazzin, a transacylase, which is the major determinant of the final acyl chain composition of the mitochondrial-specific phospholipid, CL. In addition to numerous frame shift and splice-site mutations, 36 missense mutations have been associated with BTHS. Previously, we established a BTHS-mutant panel in the yeast Saccharomyces cerevisiae that successfully models 18/21 conserved pathogenic missense mutations and defined the loss-of-function mechanism associated with a subset of the mutant tafazzins. Here, we report the biochemical and cell biological characterization of the rest of the yeast BTHS-mutant panel and in so doing identify three additional modes of tafazzin dysfunction. The largest group of mutant tafazzins is catalytically null, two mutants encode hypomorphic alleles, and another two mutants are temperature sensitive. Additionally, we have expanded the defects associated with previously characterized matrix-mislocalized-mutant tafazzins to include the rapid degradation of aggregation-prone polypeptides that correctly localize to the mitochondrial IMS. In sum, our in-depth characterization of the yeast BTHS-mutant panel has identified seven functional classes of BTHS mutation.
Figures

Similar articles
-
Barth syndrome mutations that cause tafazzin complex lability.J Cell Biol. 2011 Feb 7;192(3):447-62. doi: 10.1083/jcb.201008177. J Cell Biol. 2011. PMID: 21300850 Free PMC article.
-
Cardiolipin function in the yeast S. cerevisiae and the lessons learned for Barth syndrome.J Inherit Metab Dis. 2022 Jan;45(1):60-71. doi: 10.1002/jimd.12447. Epub 2021 Oct 19. J Inherit Metab Dis. 2022. PMID: 34626131 Free PMC article. Review.
-
Expression of human monolysocardiolipin acyltransferase-1 improves mitochondrial function in Barth syndrome lymphoblasts.J Biol Chem. 2018 May 18;293(20):7564-7577. doi: 10.1074/jbc.RA117.001024. Epub 2018 Mar 21. J Biol Chem. 2018. PMID: 29563154 Free PMC article.
-
Defining functional classes of Barth syndrome mutation in humans.Hum Mol Genet. 2016 May 1;25(9):1754-70. doi: 10.1093/hmg/ddw046. Epub 2016 Feb 16. Hum Mol Genet. 2016. PMID: 26908608 Free PMC article.
-
Barth Syndrome Cardiomyopathy: An Update.Genes (Basel). 2022 Apr 8;13(4):656. doi: 10.3390/genes13040656. Genes (Basel). 2022. PMID: 35456462 Free PMC article. Review.
Cited by
-
Single Cell Transcriptomic Analysis in a Mouse Model of Barth Syndrome Reveals Cell-Specific Alterations in Gene Expression and Intercellular Communication.Int J Mol Sci. 2023 Jul 18;24(14):11594. doi: 10.3390/ijms241411594. Int J Mol Sci. 2023. PMID: 37511352 Free PMC article.
-
Case report: Variability in clinical features as a potential pitfall for the diagnosis of Barth syndrome.Front Pediatr. 2023 Aug 16;11:1250772. doi: 10.3389/fped.2023.1250772. eCollection 2023. Front Pediatr. 2023. PMID: 37654687 Free PMC article.
-
Neutropenia in the newborn.Curr Opin Hematol. 2014 Jan;21(1):43-9. doi: 10.1097/MOH.0000000000000010. Curr Opin Hematol. 2014. PMID: 24322487 Free PMC article. Review.
-
Lipidomics Characterization of Biosynthetic and Remodeling Pathways of Cardiolipins in Genetically and Nutritionally Manipulated Yeast Cells.ACS Chem Biol. 2017 Jan 20;12(1):265-281. doi: 10.1021/acschembio.6b00995. Epub 2016 Dec 16. ACS Chem Biol. 2017. PMID: 27982579 Free PMC article.
-
Phosphatidylserine decarboxylase 1 autocatalysis and function does not require a mitochondrial-specific factor.J Biol Chem. 2015 May 15;290(20):12744-52. doi: 10.1074/jbc.M115.641118. Epub 2015 Mar 31. J Biol Chem. 2015. PMID: 25829489 Free PMC article.
References
-
- Claypool S.M., Koehler C.M. The complexity of cardiolipin in health and disease. Trends Biochem. Sci. 2012;37:32–41. doi:10.1016/j.tibs.2011.09.003. - DOI - PMC - PubMed
-
- Osman C., Voelker D.R., Langer T. Making heads or tails of phospholipids in mitochondria. J. Cell Biol. 2011;192:7–16. doi:10.1083/jcb.201006159. - DOI - PMC - PubMed
-
- Schlame M., Rua D., Greenberg M.L. The biosynthesis and functional role of cardiolipin. Prog. Lipid Res. 2000;39:257–288. doi:10.1016/S0163-7827(00)00005-9. - DOI - PubMed
-
- Dzugasova V., Obernauerova M., Horvathova K., Vachova M., Zakova M., Subik J. Phosphatidylglycerolphosphate synthase encoded by the PEL1/PGS1 gene in Saccharomyces cerevisiae is localized in mitochondria and its expression is regulated by phospholipid precursors. Curr. Genet. 1998;34:297–302. doi:10.1007/s002940050399. - DOI - PubMed
-
- Osman C., Haag M., Wieland F.T., Brugger B., Langer T. A mitochondrial phosphatase required for cardiolipin biosynthesis: the PGP phosphatase Gep4. EMBO J. 2010;29:1976–1987. doi:10.1038/emboj.2010.98. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
