

Molecular basis for SNX-BAR-mediated assembly of distinct endosomal sorting tubules
- PMID: 23085988
- PMCID: PMC3512392
- DOI: 10.1038/emboj.2012.283
Molecular basis for SNX-BAR-mediated assembly of distinct endosomal sorting tubules
Abstract
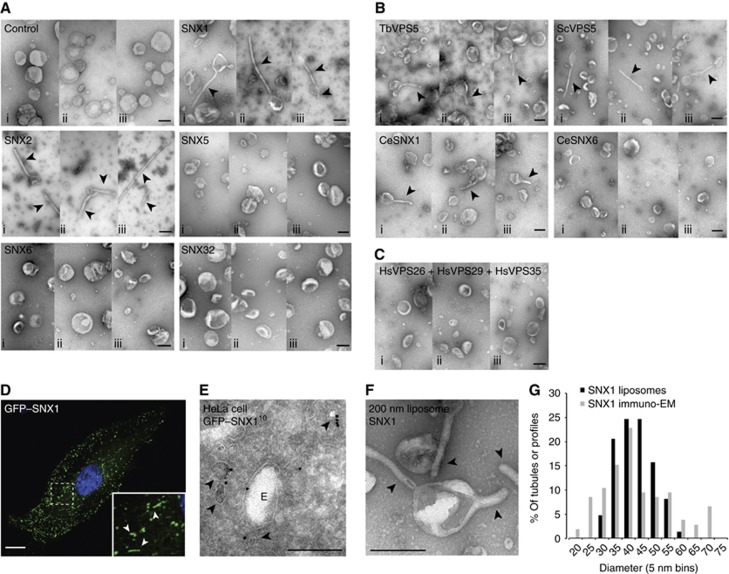
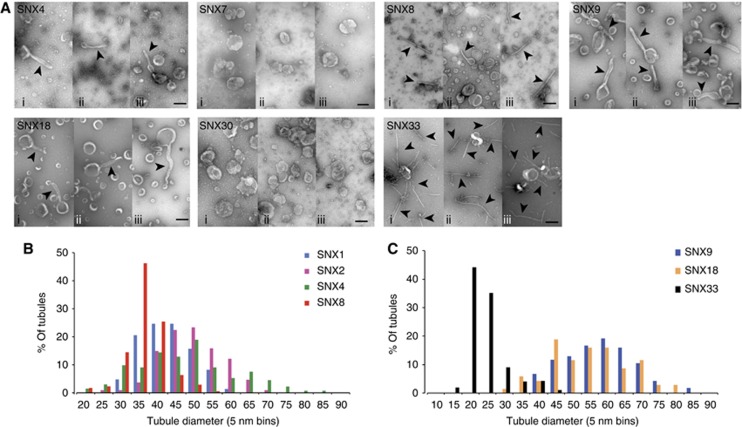
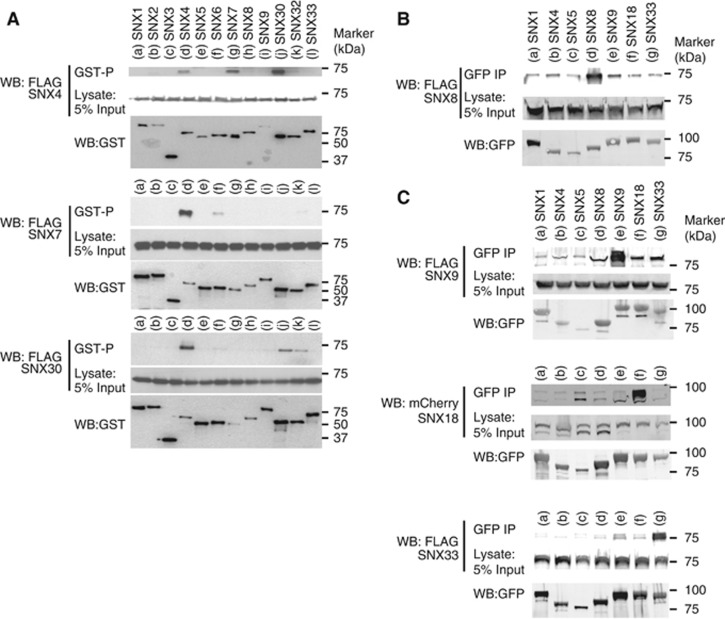
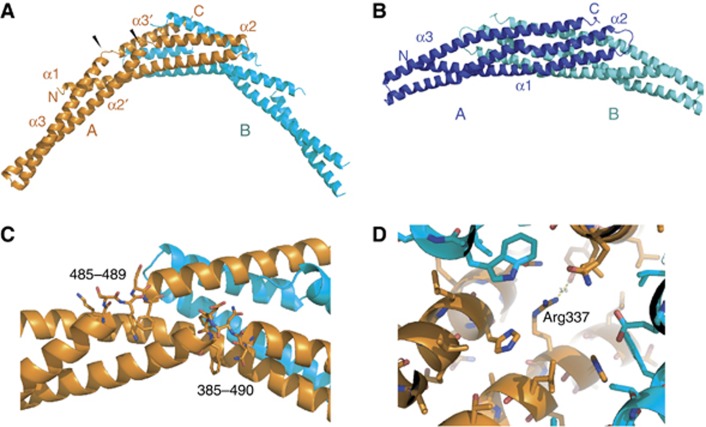
Sorting nexins (SNXs) are regulators of endosomal sorting. For the SNX-BAR subgroup, a Bin/Amphiphysin/Rvs (BAR) domain is vital for formation/stabilization of tubular subdomains that mediate cargo recycling. Here, by analysing the in vitro membrane remodelling properties of all 12 human SNX-BARs, we report that some, but not all, can elicit the formation of tubules with diameters that resemble sorting tubules observed in cells. We reveal that SNX-BARs display a restricted pattern of BAR domain-mediated dimerization, and by resolving a 2.8 Å structure of a SNX1-BAR domain homodimer, establish that dimerization is achieved in part through neutralization of charged residues in the hydrophobic BAR-dimerization interface. Membrane remodelling also requires functional amphipathic helices, predicted to be present in all SNX-BARs, and the formation of high order SNX-BAR oligomers through selective 'tip-loop' interactions. Overall, the restricted and selective nature of these interactions provide a molecular explanation for how distinct SNX-BAR-decorated tubules are nucleated from the same endosomal vacuole, as observed in living cells. Our data provide insight into the molecular mechanism that generates and organizes the tubular endosomal network.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
SNX-BAR-mediated endosome tubulation is co-ordinated with endosome maturation.Traffic. 2012 Jan;13(1):94-107. doi: 10.1111/j.1600-0854.2011.01297.x. Epub 2011 Oct 31. Traffic. 2012. PMID: 21973056
-
SNX-3 mediates retromer-independent tubular endosomal recycling by opposing EEA-1-facilitated trafficking.PLoS Genet. 2021 Jun 3;17(6):e1009607. doi: 10.1371/journal.pgen.1009607. eCollection 2021 Jun. PLoS Genet. 2021. PMID: 34081703 Free PMC article.
-
Cargo-selective SNX-BAR proteins mediate retromer trimer independent retrograde transport.J Cell Biol. 2017 Nov 6;216(11):3677-3693. doi: 10.1083/jcb.201702137. Epub 2017 Sep 21. J Cell Biol. 2017. PMID: 28935632 Free PMC article.
-
You can go your own way: SNX-BAR coat complexes direct traffic at late endosomes.Curr Opin Cell Biol. 2022 Jun;76:102087. doi: 10.1016/j.ceb.2022.102087. Epub 2022 May 12. Curr Opin Cell Biol. 2022. PMID: 35569261 Review.
-
SNX-BAR proteins in phosphoinositide-mediated, tubular-based endosomal sorting.Semin Cell Dev Biol. 2010 Jun;21(4):371-80. doi: 10.1016/j.semcdb.2009.11.009. Epub 2009 Nov 13. Semin Cell Dev Biol. 2010. PMID: 19914387 Free PMC article. Review.
Cited by
-
The phosphorylation of sorting nexin 5 at serine 226 regulates retrograde transport and macropinocytosis.PLoS One. 2018 Nov 12;13(11):e0207205. doi: 10.1371/journal.pone.0207205. eCollection 2018. PLoS One. 2018. PMID: 30419003 Free PMC article.
-
CROP: a retromer-PROPPIN complex mediating membrane fission in the endo-lysosomal system.EMBO J. 2022 May 16;41(10):e109646. doi: 10.15252/embj.2021109646. Epub 2022 Apr 25. EMBO J. 2022. PMID: 35466426 Free PMC article.
-
ER morphology and endo-lysosomal crosstalk: Functions and disease implications.Biochim Biophys Acta Mol Cell Biol Lipids. 2020 Jan;1865(1):158544. doi: 10.1016/j.bbalip.2019.158544. Epub 2019 Oct 31. Biochim Biophys Acta Mol Cell Biol Lipids. 2020. PMID: 31678515 Free PMC article. Review.
-
Protein sorting from endosomes to the TGN.Front Cell Dev Biol. 2023 Feb 21;11:1140605. doi: 10.3389/fcell.2023.1140605. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 36895788 Free PMC article. Review.
-
Nuclear epidermal growth factor receptor as a therapeutic target.Explor Target Antitumor Ther. 2023;4(4):616-629. doi: 10.37349/etat.2023.00156. Epub 2023 Aug 30. Explor Target Antitumor Ther. 2023. PMID: 37720348 Free PMC article. Review.
References
-
- Blanc E, Roversi P, Vonrhein C, Flensburg C, Lea SM, Bricogne G (2004) Refinement of severely incomplete structures with maximum likelihood in BUSTER-TNT. Acta Crystallogr D Biol Crystallogr 60: 2210–2221 - PubMed
-
- Bujny MV, Popoff V, Johannes L, Cullen PJ (2007) The retromer component sorting nexin-1 is required for efficient retrograde transport of Shiga toxin from early endosome to the trans Golgi network. J Cell Sci 120: 2010–2021 - PubMed
-
- Carlton J, Bujny M, Peter B, Oorschot V, Rutherford A, Mellor H, Klumperman J, McMahon HT, Cullen PJ (2004) Sorting nexin-1 mediates tubular endosome-to-TGN transport through coincidence sensing of high- curvature membranes and 3-phosphoinositides. Curr Biol 14: 1791–1800 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
