

Review
doi: 10.1016/B978-0-12-404742-6.00002-8.
Precision editing of large animal genomes
Affiliations
- PMID: 23084873
- PMCID: PMC3683964
- DOI: 10.1016/B978-0-12-404742-6.00002-8
Item in Clipboard
Review
Precision editing of large animal genomes
Adv Genet.
2012.
Abstract
Transgenic animals are an important source of protein and nutrition for most humans and will play key roles in satisfying the increasing demand for food in an ever-increasing world population. The past decade has experienced a revolution in the development of methods that permit the introduction of specific alterations to complex genomes. This precision will enhance genome-based improvement of farm animals for food production. Precision genetics also will enhance the development of therapeutic biomaterials and models of human disease as resources for the development of advanced patient therapies.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures

The multiple applications of genetically modified large animals. The pig is shown as an example. The first application is to improve traits in the farm animal. Examples of the potential improved traits include (1) resistance to diseases, (2) improved nutrition such as introducing a gene to produce the healthier omega-3 fatty acids to replace the normal omega-6 fatty acids (Lai et al., 2006), and (3) reducing the environmental impact of major pig production facilities by reducing phosphorous in manure (Golovan et al., 2001). The second application of genetically modified pigs is for biomedical products such as organ transplantation (http://web.archive.org/web/20071210031618/http://www.fda.gov/fdac/features/596_xeno.html ) or specific functional organ parts such as heart valves and subcellular structures. Examples include inactivating genes such as α-1,3-galactose that produce powerful immune responses when introduced into humans and eliminating the potential spread of porcine endogenous retroviruses. The third application of genetically modified pigs is the creation of animals that closely mimic human diseases such as cystic fibrosis (Rogers et al., 2008), cardiovascular disease, and cancer. For color version of this figure, the reader is referred to the online version of this book.
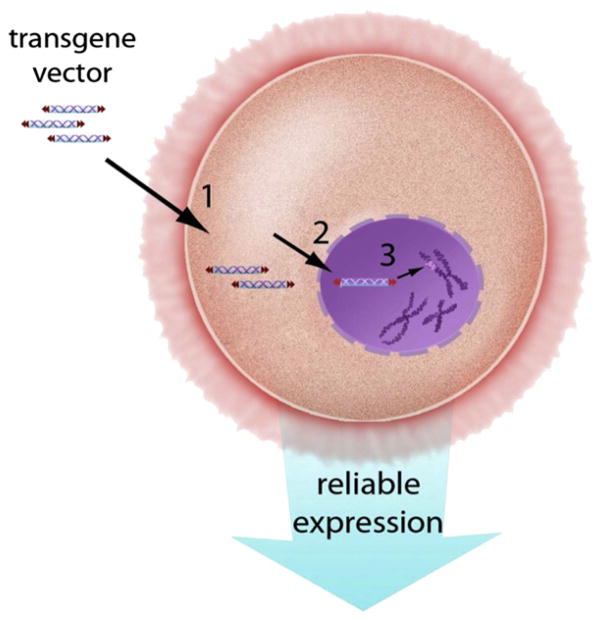
The three barriers to the introduction of foreign DNA into genomes: (1) the cell membrane, (2) the nuclear membrane, and (3) the chromosomal DNA in the chromosomes. For effective transgenesis, the foreign DNA must overcome the three barriers and then be able to withstand protective measures such as methylation that are employed to reduce expression of transgenic DNA that has inserted into the chromatin. For color version of this figure, the reader is referred to the online version of this book.
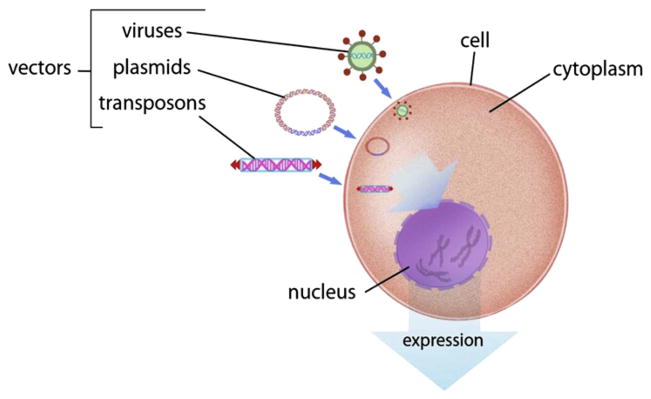
The three vectors for introduction of foreign DNA into genomes: (1) plasmids, (2) viruses, and (3) transposons. For color version of this figure, the reader is referred to the online version of this book.
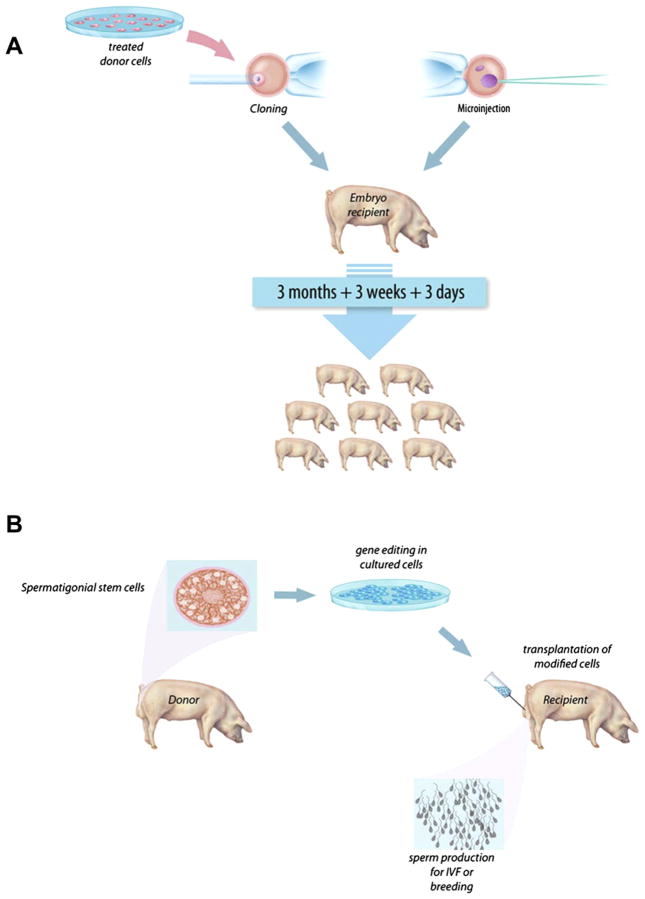
Methods for genetic modification in livestock. (A) A flow diagram of the primary steps involved with the production of transgenic livestock by SCNT (cloning) and embryo microinjection. For simplicity, the illustrations show pigs only, but the general procedure applies to each of the major livestock species. Each procedure requires either surgical or in vitro production of oocytes or embryos. Donor cells used for SCNT (left) can be genetically modified in culture by a number of methods described in this review. Modified donor cells are injected into enucleated oocytes, which are then fused and activated prior to embryo transfer into a recipient. Embryo microinjection (right) is performed on zygotes 18–24 h after fertilization. The injection site can vary, but typically, DNA is delivered directly to the pronucleus by pronuclear injection, SB trans-posons plus transposase mRNA, ZFN, or TALEN mRNA can be injected into the cytoplasm, and viral particles are typically injected into the perivitelline space. Embryos manipulated in each case are implanted into a synchronized recipient female to establish pregnancy. Resulting offspring can be screened for the desired modifications and expression patterns. (B) Spermatogonial stem cells offer a second method for genetic modification of livestock. Genetic modification or gene edits can be executed in vitro in spermatogonial stem cells isolated from donor testes. Modified cells are transplanted into germ cell-depleted testes of a recipient. Implanted spermatogonial stem cells produce sperm that carry the genetic modification(s) that can be used for breeding via artificial insemination or in vitro fertilization (IVF) to derive founder animals. For color version of this figure, the reader is referred to the online version of this book.
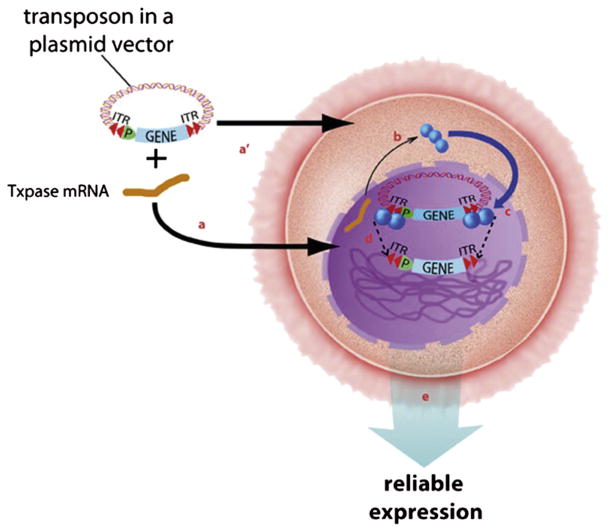
DNA transposition consists of an enzymatic cut-and-paste reaction in which a transposon containing a gene of interest [shown in blue, with its promoter (P)] is cut out of a plasmid and inserted into a chromosome. The cleavage reaction occurs at the ends of the ITRs (inverted set of red double arrowheads) of the transposon. The transposons integrate only into TA-dinucleotide basepairs (about 200 million in a mammalian genome). The ITRs are the only DNA sequences required by the transposase enzyme for transposition. The transposase enzyme (Txpase, blue balls) drives the cut-and-paste reaction. Transposase activity is obtained by co-injecting transposon and an mRNA encoding the Txpase (blue squiggle) into either the nucleus (a) or cytoplasm (a′). The plasmid carrying the transposon and transposase-encoding mRNA enter a cell (large back oval) and proceed through the nuclear membrane (dashed line) (b). The transposase mRNA is translated in the cytoplasm to give an appropriate level of enzyme (c). The transposase molecules enter the nucleus and bind to the transposon, two at each end (c). Four transposase enzymes work in concert to cleave the plasmid at the termini of the transposon and paste it (dotted lines) into chromosomal DNA (green tangled lines) (e). Monomeric integration into a chromosome can confer reliable expression of the gene of interest that is contained within the transposon through multiple generations. For color version of this figure, the reader is referred to the online version of this book.
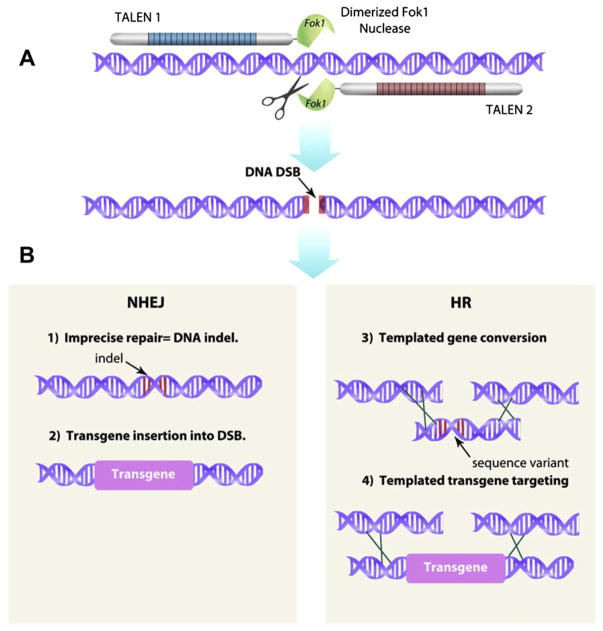
Site-specific targeting of genetic changes using hybrid DNases. (A) A pair of TALEN nucleases is shown as an example of hybrid DNases designed to cleave at a unique sequence in a genome. The pair of TALENs executes a double-strand DNA break (DSB) at the targeted locus. (B) If no other DNA sequences are added, the DSB will be repaired by the process of NHEJ that will generally result in a minor insertion or deletion of a few basepairs (indels; example 1). Alternatively, because the NHEJ DNA repair enzymes that assemble at the DSB can facilitate the integration of a foreign DNA sequence, a transgene can be introduced into the site with higher than random efficiency (example 2). Alternatively, if a DNA sequence that has a high identity with the region surrounding the DSB is introduced, homologous recombination (HR) can occur (examples 3 and 4). The introduced DNA sequence may vary by only a single (or a few) basepair, which results in a defined mutation that is equivalent to a natural allele (example 3). However, if an entire expression cassette with a foreign transgene is flanked by homologous sequences at the DSB, then the transgene will have a high probability of being copied precisely into the DSB (example 4). For color version of this figure, the reader is referred to the online version of this book.
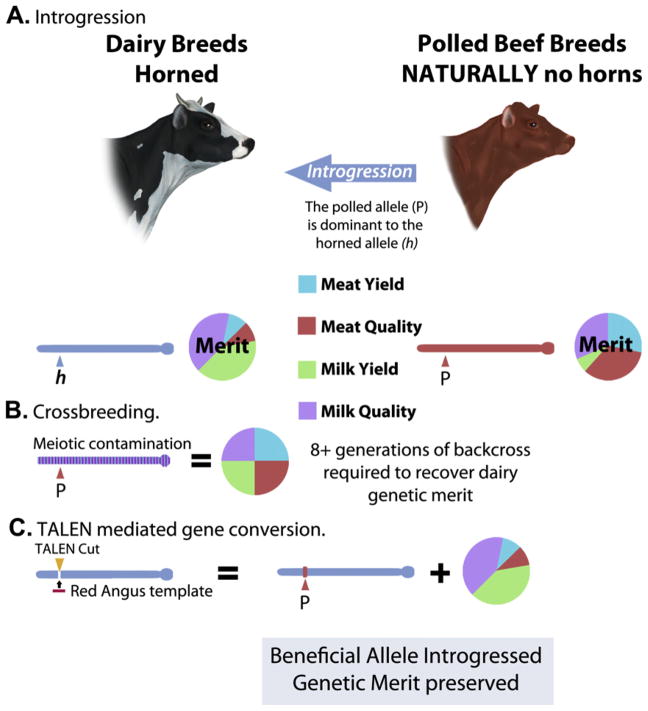
A) The diagram contrasts introgression of desired alleles (polled allele to horned animals) by crossbreeding (panel B) versus TALEN-mediated gene conversion (panel C). Beef and dairy breeds are selected for divergent classes of traits resulting in genetic merit selected for production of meat or milk, respectively. The accumulation of these traits is referred to as the genetic merit of each animal. Crossbreeding mixes these traits, which would result in animals that would not be ideal for either milk or meat production. The trait-selected genome architecture of these animals is conflicted by meiotic contamination, which would require about eight generations of selection to recover the original genetic merit. Panel C shows how TALEN-mediated gene conversion is able to transfer just a desired trait from beef cattle into dairy breeds. In this example, TALENs generate a double-strand DNA break at the horned-polled locus that can be repaired by a homologous template carrying the polled allele from a polled beef breed, e.g., Red Angus. The resulting animal will be both free of horns and maintain the original genetic architecture and merit for milk production. For color version of this figure, the reader is referred to the online version of this book.
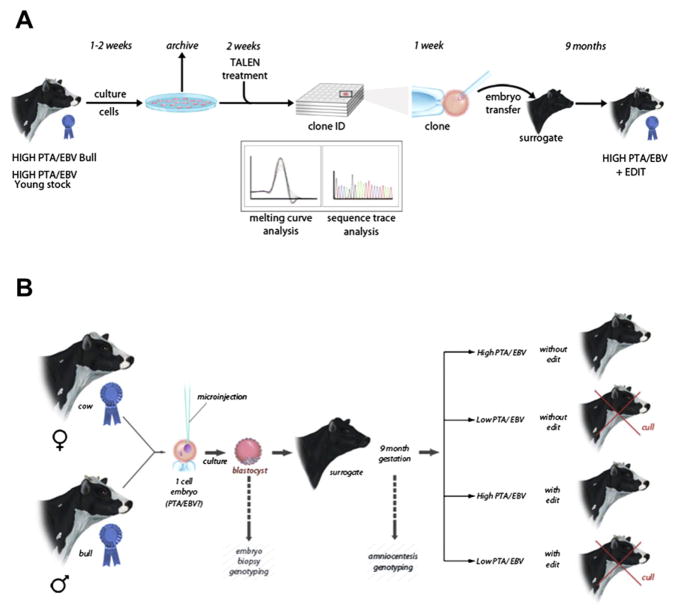
Strategies for implementation of allelic introgression. The introgression of desirable alleles into livestock could follow either a vertical (panel A) or a horizontal (panel B) paradigm. (A) In the vertical paradigm, allelic introgression would be performed in cells derived from a donor individual(s) with a high predictability of transmitting ability/estimated breeding value (PTA/EBV, denoted by a blue ribbon). One or several genetic heterozygous or homozygous allele conversions (genetic edits) could be made and verified (e.g., by sequence analysis) prior to cloning of an individual. The resulting animal would not only carry the edits but would also maintain the original PTA/EBV of the donor animal. This animal would be entered back into the genetic improvement program and edits would be selected in subsequent generations. (B) Horizontal implementation takes advantage of the fact that embryo transfer is routine in genetic improvement programs of some livestock species, e.g., cattle. Zygotes produced from animals with high PTA/EBV could be injected with TALENs plus repair templates corresponding to the desired alleles and implanted into a surrogate for establishment or pregnancy. Resulting offspring could be scored for high PTA/EBV and either the presence or the absence of the targeted edits. Animals with high PTA/EBV would be maintained in the genetic improvement program regardless of the edit status, while animals with low PTA/EBV would be culled. Two potential improvements of this process can be envisioned. (1) An embryo biopsy at the blastocysts stage could be collected to evaluate the edit status or PTA/EBV so that only edited and/or high PTA/EBV embryos would be implanted into surrogates. (2) Fetal cells could be collected early in pregnancy by amniocentesis for evaluation of the edit status or PTA/ EBV. Low PTA/EBV or non-edited animals could be culled prior to parturition. Development of these technologies could further accelerate the rate of livestock improvement. In contrast to the vertical paradigm, allelic introgression and genetic improvement will continue to occur in the horizontal paradigm, thereby producing animals that would be one generation ahead in terms of genetic improvement. This method could be easily applied to generate numerous animals from multiple lines such that dissemination of converted alleles (genetic edits) would be accomplished rapidly within a population with minimal risk of inbreeding. For color version of this figure, the reader is referred to the online version of this book.
Similar articles
-
Precision genetics for complex objectives in animal agriculture.J Anim Sci. 2010 Jul;88(7):2530-9. doi: 10.2527/jas.2010-2847. Epub 2010 Mar 12. J Anim Sci. 2010. PMID: 20228236 Free PMC article.
-
Transgenic farm animals: applications in agriculture and biomedicine.Biotechnol Annu Rev. 2000;5:269-92. doi: 10.1016/s1387-2656(00)05039-0. Biotechnol Annu Rev. 2000. PMID: 10875004 Review.
-
Genome editing for crop improvement: Challenges and opportunities.GM Crops Food. 2015;6(4):183-205. doi: 10.1080/21645698.2015.1129937. GM Crops Food. 2015. PMID: 26930114 Free PMC article.
-
How farm animals are improving human health and welfare.Rev Sci Tech. 2018 Apr;37(1):83-96. doi: 10.20506/rst.37.1.2742. Rev Sci Tech. 2018. PMID: 30209427 Review.
-
[Application of CRISPR/Cas9 mediated genome editing in farm animals].Yi Chuan. 2016 Mar;38(3):217-26. doi: 10.16288/j.yczz.15-398. Yi Chuan. 2016. PMID: 27001476 Review. Chinese.
Cited by
-
Model organisms: beyond the inner circle.Nat Methods. 2013 Jun;10(6):471-3. doi: 10.1038/nmeth.2484. Nat Methods. 2013. PMID: 23722204 No abstract available.
-
Exogenous enzymes upgrade transgenesis and genetic engineering of farm animals.Cell Mol Life Sci. 2015 May;72(10):1907-29. doi: 10.1007/s00018-015-1842-1. Epub 2015 Feb 1. Cell Mol Life Sci. 2015. PMID: 25636347 Free PMC article. Review.
-
CRISPR-Based Genome Editing as a New Therapeutic Tool in Retinal Diseases.Mol Biotechnol. 2021 Sep;63(9):768-779. doi: 10.1007/s12033-021-00345-4. Epub 2021 May 31. Mol Biotechnol. 2021. PMID: 34057656 Review.
-
Is gene editing an acceptable alternative to castration in pigs?PLoS One. 2019 Jun 24;14(6):e0218176. doi: 10.1371/journal.pone.0218176. eCollection 2019. PLoS One. 2019. PMID: 31233520 Free PMC article.
-
Live pigs produced from genome edited zygotes.Sci Rep. 2013 Oct 10;3:2847. doi: 10.1038/srep02847. Sci Rep. 2013. PMID: 24108318 Free PMC article.
References
-
- Arnould S, Delenda C, Grizot S, Desseaux C, Pâques F, Silva GH, et al. The I-CreI meganuclease and its engineered derivatives: applications from cell modification to gene therapy. Protein Engineering Design and Selection. 2011;24:27–31. - PubMed
-
- Baguisi A, Behboodi E, Melican DT, Pollock JS, Destrempes MM, Cammuso C, et al. Production of goats by somatic cell nuclear transfer. Nature Biotechnology. 1999;17:456–461. - PubMed
-
- Baldassarre H, Wang B, Kafidi N, Gauthier M, Neveu N, Lapointe J, et al. Production of transgenic goats by pronuclear microinjection of in vitro produced zygotes derived from oocytes recovered by laparoscopy. Theriogenology. 2003;59:831–839. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
