

Bacterial neuraminidase rescues influenza virus replication from inhibition by a neuraminidase inhibitor
- PMID: 23028967
- PMCID: PMC3445474
- DOI: 10.1371/journal.pone.0045371
Bacterial neuraminidase rescues influenza virus replication from inhibition by a neuraminidase inhibitor
Abstract
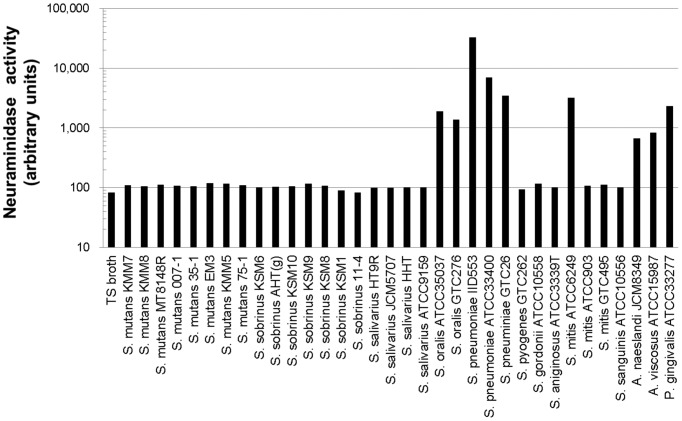
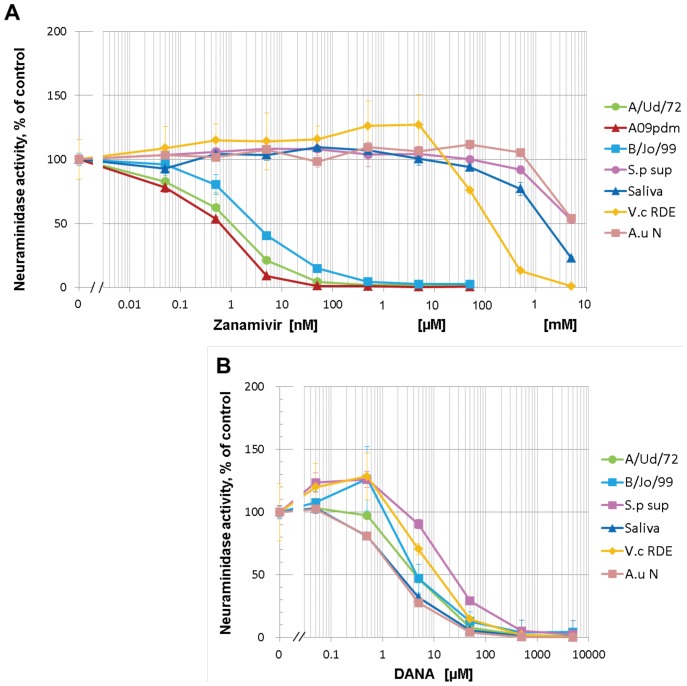
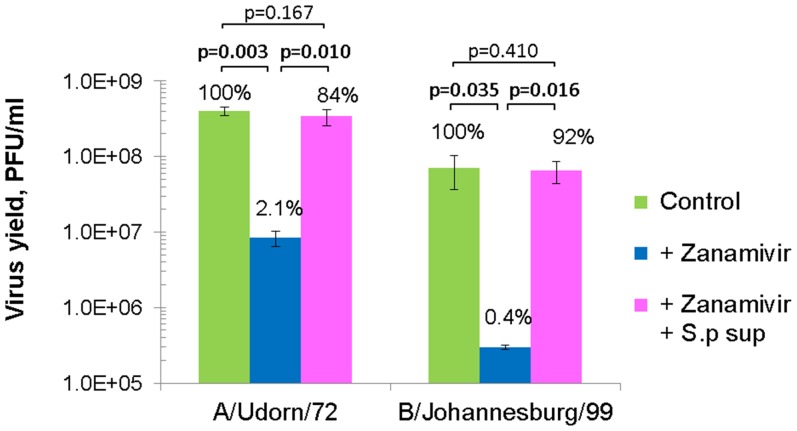
Influenza virus neuraminidase (NA) cleaves terminal sialic acid residues on oligosaccharide chains that are receptors for virus binding, thus playing an important role in the release of virions from infected cells to promote the spread of cell-to-cell infection. In addition, NA plays a role at the initial stage of viral infection in the respiratory tract by degrading hemagglutination inhibitors in body fluid which competitively inhibit receptor binding of the virus. Current first line anti-influenza drugs are viral NA-specific inhibitors, which do not inhibit bacterial neuraminidases. Since neuraminidase producing bacteria have been isolated from oral and upper respiratory commensal bacterial flora, we posited that bacterial neuraminidases could decrease the antiviral effectiveness of NA inhibitor drugs in respiratory organs when viral NA is inhibited. Using in vitro models of infection, we aimed to clarify the effects of bacterial neuraminidases on influenza virus infection in the presence of the NA inhibitor drug zanamivir. We found that zanamivir reduced progeny virus yield to less than 2% of that in its absence, however the yield was restored almost entirely by the exogenous addition of bacterial neuraminidase from Streptococcus pneumoniae. Furthermore, cell-to-cell infection was severely inhibited by zanamivir but restored by the addition of bacterial neuraminidase. Next we examined the effects of bacterial neuraminidase on hemagglutination inhibition and infectivity neutralization activities of human saliva in the presence of zanamivir. We found that the drug enhanced both inhibitory activities of saliva, while the addition of bacterial neuraminidase diminished this enhancement. Altogether, our results showed that bacterial neuraminidases functioned as the predominant NA when viral NA was inhibited to promote the spread of infection and to inactivate the neutralization activity of saliva. We propose that neuraminidase from bacterial flora in patients may reduce the efficacy of NA inhibitor drugs during influenza virus infection. (295 words).
Conflict of interest statement
Figures
Similar articles
-
Resistance to Mutant Group 2 Influenza Virus Neuraminidases of an Oseltamivir-Zanamivir Hybrid Inhibitor.J Virol. 2016 Nov 14;90(23):10693-10700. doi: 10.1128/JVI.01703-16. Print 2016 Dec 1. J Virol. 2016. PMID: 27654293 Free PMC article.
-
Neuraminidase inhibitor drug susceptibility differs between influenza N1 and N2 neuraminidase following mutagenesis of two conserved residues.Antiviral Res. 2007 Dec;76(3):263-6. doi: 10.1016/j.antiviral.2007.08.002. Epub 2007 Aug 27. Antiviral Res. 2007. PMID: 17868928
-
Effect of hemagglutinin glycosylation on influenza virus susceptibility to neuraminidase inhibitors.J Virol. 2005 Oct;79(19):12416-24. doi: 10.1128/JVI.79.19.12416-12424.2005. J Virol. 2005. PMID: 16160169 Free PMC article.
-
Neuraminidase inhibitors as antiviral agents.Curr Drug Targets Infect Disord. 2005 Dec;5(4):401-9. doi: 10.2174/156800505774912884. Curr Drug Targets Infect Disord. 2005. PMID: 16535861 Review.
-
Anti-influenza drugs and neuraminidase inhibitors.Prog Drug Res. 2001;Spec No:35-77. doi: 10.1007/978-3-0348-7784-8_2. Prog Drug Res. 2001. PMID: 11548210 Review.
Cited by
-
A cooperativity between virus and bacteria during respiratory infections.Front Microbiol. 2023 Nov 30;14:1279159. doi: 10.3389/fmicb.2023.1279159. eCollection 2023. Front Microbiol. 2023. PMID: 38098657 Free PMC article. Review.
-
Using common spatial distributions of atoms to relate functionally divergent influenza virus N10 and N11 protein structures to functionally characterized neuraminidase structures, toxin cell entry domains, and non-influenza virus cell entry domains.PLoS One. 2015 Feb 23;10(2):e0117499. doi: 10.1371/journal.pone.0117499. eCollection 2015. PLoS One. 2015. PMID: 25706124 Free PMC article.
-
Capsular sialic acid of Streptococcus suis serotype 2 binds to swine influenza virus and enhances bacterial interactions with virus-infected tracheal epithelial cells.Infect Immun. 2013 Dec;81(12):4498-508. doi: 10.1128/IAI.00818-13. Epub 2013 Sep 30. Infect Immun. 2013. PMID: 24082069 Free PMC article.
-
Influenza Therapeutics in Clinical Practice-Challenges and Recent Advances.Cold Spring Harb Perspect Med. 2021 Apr 1;11(4):a038463. doi: 10.1101/cshperspect.a038463. Cold Spring Harb Perspect Med. 2021. PMID: 32041763 Free PMC article. Review.
-
Inhibitory effects of Patchouli alcohol on the early lifecycle stages of influenza A virus.Front Microbiol. 2023 Feb 1;13:938868. doi: 10.3389/fmicb.2022.938868. eCollection 2022. Front Microbiol. 2023. PMID: 36817112 Free PMC article.
References
-
- von Itzstein M, Wu WY, Kok GB, Pegg MS, Dyason JC, et al. (1993) Rational design of potent sialidase-based inhibitors of influenza virus replication. Nature 363: 418–423. - PubMed
-
- Palese P, Shaw ML (2007) Orthomixoviride: The virus replication. Knipe DM, Howley PM, editors. Philadelphia: Lippincotto Williams & Wilkins.
-
- Palese P, Compans RW (1976) Inhibition of influenza virus replication in tissue culture by 2-deoxy-2,3-dehydro-N-trifluoroacetylneuraminic acid (FANA): mechanism of action. J Gen Virol 33: 159–163. - PubMed
-
- Boat TF, Davis J, Stern RC, Cheng PW (1978) Effect of blood group determinants on binding of human salivary mucous glycoproteins to influenza virus. Biochim Biophys Acta 540: 127–133. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
