

bHLH-PAS heterodimer of methoprene-tolerant and Cycle mediates circadian expression of juvenile hormone-induced mosquito genes
- PMID: 23012454
- PMCID: PMC3478602
- DOI: 10.1073/pnas.1214209109
bHLH-PAS heterodimer of methoprene-tolerant and Cycle mediates circadian expression of juvenile hormone-induced mosquito genes
Abstract
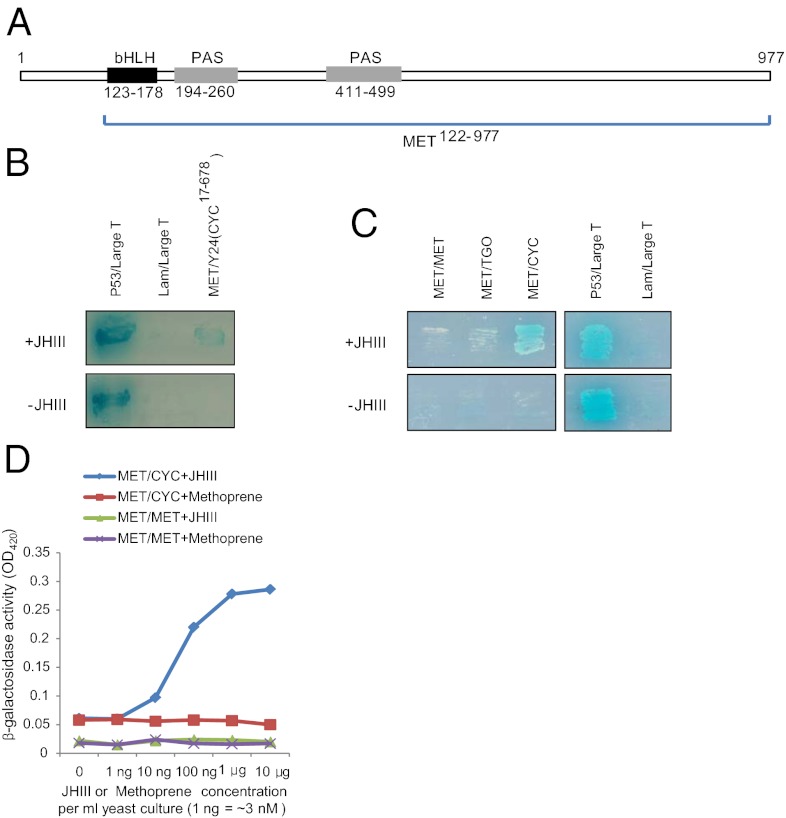
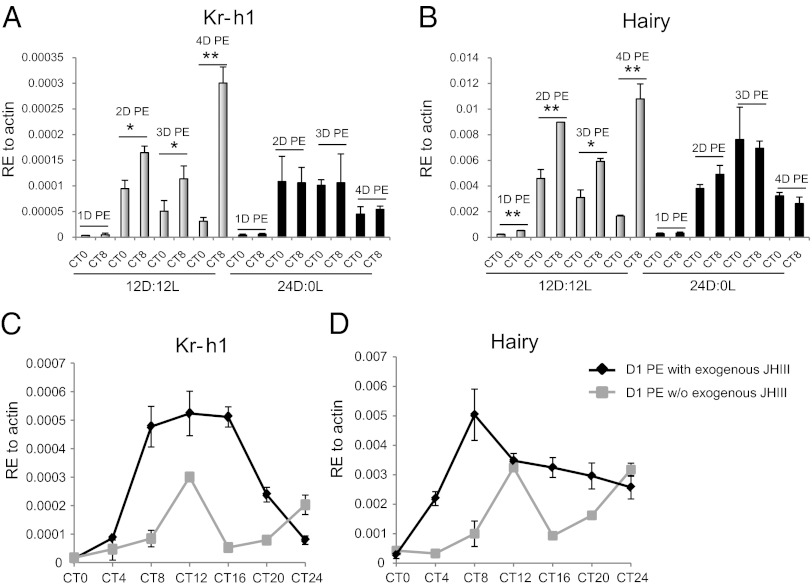
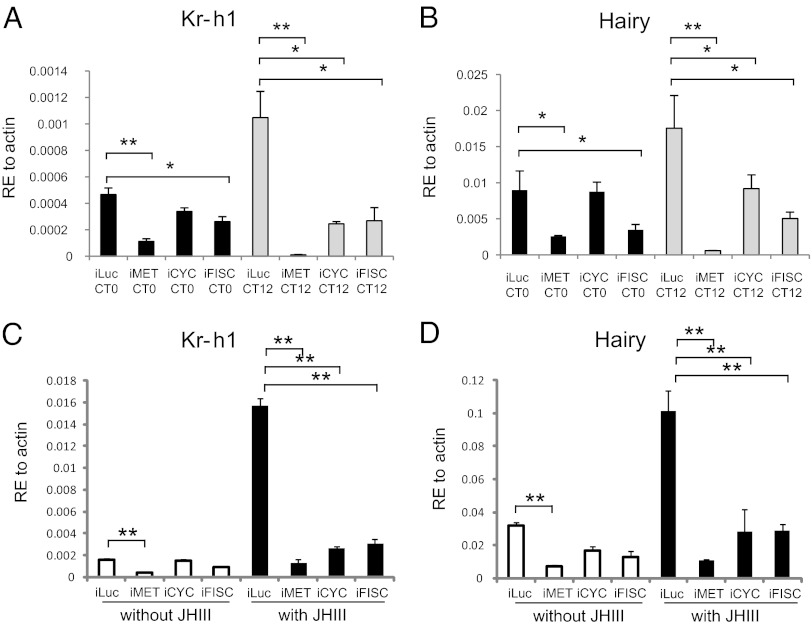
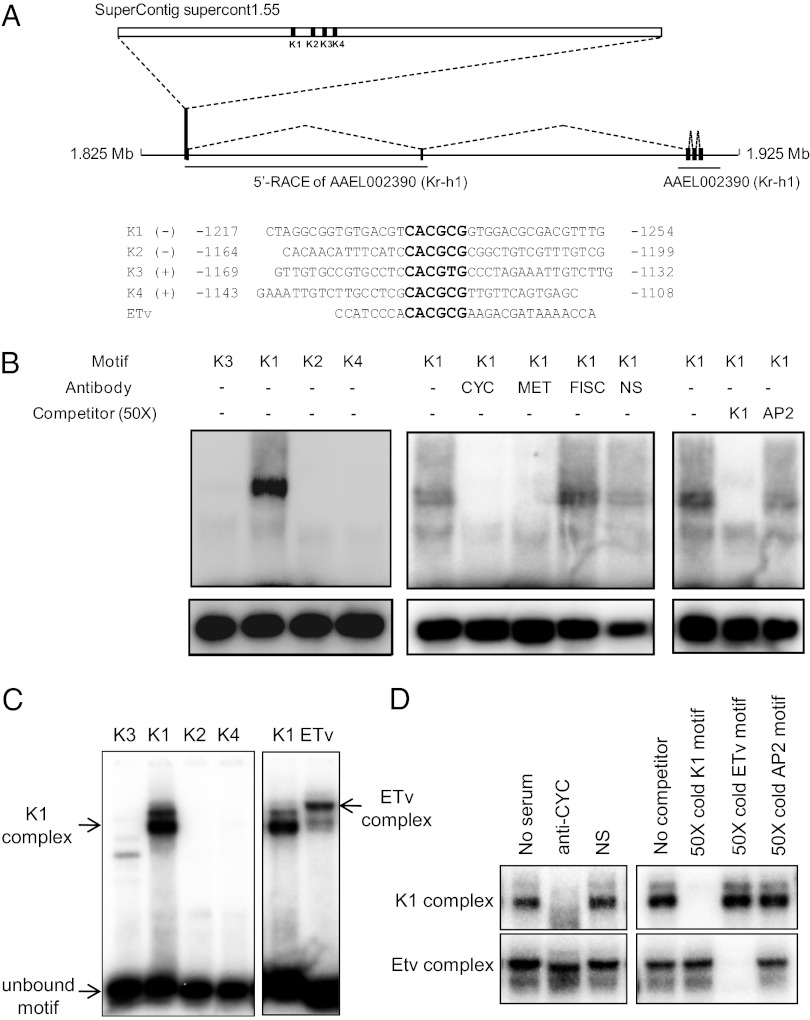
Juvenile hormone (JH) governs a great diversity of processes in insect development and reproduction. It plays a critical role in controlling the gonadotrophic cycles of female mosquitoes by preparing tissues for blood digestion and egg development. Here, we show that in female Aedes aegypti mosquitoes JH III control of gene expression is mediated by a heterodimer of two bHLH-PAS proteins-the JH receptor methoprene-tolerant (MET) and Cycle (CYC, AAEL002049). We identified Aedes CYC as a MET-interacting protein using yeast two-hybrid screening. Binding of CYC and MET required the presence of JH III. In newly eclosed female mosquitoes, the expression of two JH-responsive genes, Kr-h1 and Hairy, was dependent on both the ratio of light to dark periods and JH III. Their expression was compromised by in vivo RNA interference (RNAi) depletions of CYC, MET, and the steroid receptor coactivator SRC/FISC. Moreover, JH III was not effective in induction of Kr-h1 and Hairy gene expression in vitro in fat bodies of female mosquitoes with RNAi-depleted CYC, MET or SRC/FISC. A sequence containing an E-box-like motif from the Aedes Kr-h1 gene promoter specifically interacted with a protein complex, which included MET and CYC from the female mosquito fat body nuclear extract. These results indicate that a MET/CYC heterodimer mediates JH III activation of Kr-h1 and Hairy genes in the context of light-dependent circadian regulation in female mosquitoes during posteclosion development. This study provides an important insight into the understanding of the molecular basis of JH action.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Synergistic action of the transcription factors Krüppel homolog 1 and Hairy in juvenile hormone/Methoprene-tolerant-mediated gene-repression in the mosquito Aedes aegypti.PLoS Genet. 2019 Oct 29;15(10):e1008443. doi: 10.1371/journal.pgen.1008443. eCollection 2019 Oct. PLoS Genet. 2019. PMID: 31661489 Free PMC article.
-
Hairy and Groucho mediate the action of juvenile hormone receptor Methoprene-tolerant in gene repression.Proc Natl Acad Sci U S A. 2016 Feb 9;113(6):E735-43. doi: 10.1073/pnas.1523838113. Epub 2016 Jan 7. Proc Natl Acad Sci U S A. 2016. PMID: 26744312 Free PMC article.
-
A steroid receptor coactivator acts as the DNA-binding partner of the methoprene-tolerant protein in regulating juvenile hormone response genes.Mol Cell Endocrinol. 2014 Aug 25;394(1-2):47-58. doi: 10.1016/j.mce.2014.06.021. Epub 2014 Jul 6. Mol Cell Endocrinol. 2014. PMID: 25004255 Free PMC article.
-
Methoprene-tolerant and Krüppel homolog 1 are actors of juvenile hormone-signaling controlling the development of male sexual behavior in the moth Agrotis ipsilon.Horm Behav. 2023 Apr;150:105330. doi: 10.1016/j.yhbeh.2023.105330. Epub 2023 Feb 13. Horm Behav. 2023. PMID: 36791650 Review.
-
Juvenile hormone signaling - a mini review.Insect Sci. 2019 Aug;26(4):600-606. doi: 10.1111/1744-7917.12614. Epub 2018 Jul 18. Insect Sci. 2019. PMID: 29888456 Review.
Cited by
-
Identification of juvenile hormone-induced posttranslational modifications of methoprene tolerant and Krüppel homolog 1 in the yellow fever mosquito, Aedes aegypti.J Proteomics. 2021 Jun 30;242:104257. doi: 10.1016/j.jprot.2021.104257. Epub 2021 May 4. J Proteomics. 2021. PMID: 33957312 Free PMC article.
-
Krüppel-homolog 1 exerts anti-metamorphic and vitellogenic functions in insects via phosphorylation-mediated recruitment of specific cofactors.BMC Biol. 2021 Oct 8;19(1):222. doi: 10.1186/s12915-021-01157-3. BMC Biol. 2021. PMID: 34625063 Free PMC article.
-
Microarrays reveal discrete phases in juvenile hormone regulation of mosquito reproduction.Proc Natl Acad Sci U S A. 2013 Jun 11;110(24):9623-4. doi: 10.1073/pnas.1307487110. Epub 2013 May 29. Proc Natl Acad Sci U S A. 2013. PMID: 23720307 Free PMC article. No abstract available.
-
Methoprene-Tolerant (Met) Is Indispensable for Larval Metamorphosis and Female Reproduction in the Cotton Bollworm Helicoverpa armigera.Front Physiol. 2018 Nov 15;9:1601. doi: 10.3389/fphys.2018.01601. eCollection 2018. Front Physiol. 2018. PMID: 30498452 Free PMC article.
-
Genome-wide analyses of the bHLH superfamily in crustaceans: reappraisal of higher-order groupings and evidence for lineage-specific duplications.R Soc Open Sci. 2018 Mar 14;5(3):172433. doi: 10.1098/rsos.172433. eCollection 2018 Mar. R Soc Open Sci. 2018. PMID: 29657824 Free PMC article.
References
-
- Clements AN. 1992. The biology of mosquitoes. Development, Nutrition and Reproduction (Chapman & Hall, London), Vol 1.
-
- Raikhel AS, et al. Molecular biology of mosquito vitellogenesis: from basic studies to genetic engineering of antipathogen immunity. Insect Biochem Mol Biol. 2002;32:1275–1286. - PubMed
-
- Gwadz RW, Spielman A. Corpus allatum control of ovarian development in Aedes aegypti. J Insect Physiol. 1973;19:1441–1448. - PubMed
-
- Raikhel AS, Lea AO. Hormone-mediated formation of the endocytic complex in mosquito oocytes. Gen Comp Endocrinol. 1985;57:422–433. - PubMed
-
- Raikhel AS, Lea AO. Juvenile hormone controls previtellogenic proliferation of ribosomal RNA in the mosquito fat body. Gen Comp Endocrinol. 1990;77:423–434. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
