

Prkar1a in the regulation of insulin secretion
- PMID: 22951902
- PMCID: PMC4034137
- DOI: 10.1055/s-0032-1321866
Prkar1a in the regulation of insulin secretion
Abstract
The incidence of type 2 diabetes mellitus (T2DM) is rapidly increasing worldwide with significant consequences on individual quality of life as well as economic burden on states' healthcare costs. While origins of the pathogenesis of T2DM are poorly understood, an early defect in glucose-stimulated insulin secretion (GSIS) from pancreatic β-cells is considered a hallmark of T2DM. Upon a glucose stimulus, insulin is secreted in a biphasic manner with an early first-phase burst of insulin, which is followed by a second, more sustained phase of insulin output. First phase insulin secretion is diminished early in T2DM as well is in subjects who are at risk of developing T2DM. An effective treatment of T2DM with incretin hormone glucagon-like peptide-1 (GLP-1) or its long acting peptide analogue exendin-4 (E4), restores first-phase and augments second-phase glucose stimulated insulin secretion. This effect of incretin action occurs within minutes of GLP-1/E4 infusion in T2DM humans. An additional important consideration is that incretin hormones augment GSIS only above a certain glucose threshold, which is slightly above the normal glucose range. This ensures that incretin hormones stimulate GSIS only when glucose levels are high, while they are ineffective when insulin levels are below a certain threshold. Activation of the GLP-1 receptor, which is highly expressed on pancreatic β-cells, stimulates 2 -distinct intracellular signaling pathways: a) the cAMP-protein kinase A branch and b) the cAMP-EPAC2 (EPAC=exchange protein activated by cAMP) branch. While the EPAC2 branch is considered to mediate GLP-1 effects on first-phase GSIS, the PKA branch is necessary for the former branch to be active. However, how these 2 branches interplay and converge and how their effects on insulin secretion and insulin vesicle exocytosis are coordinated is poorly understood.Thus, at the outset of our studies we have a poorly understood intracellular interplay of cAMP-dependent signaling pathways, which - when stimulated - restore glucose-dependent first phase and augment second phase insulin secretion in the ailing β-cells of T2DM.
© Georg Thieme Verlag KG Stuttgart · New York.
Conflict of interest statement
The authors have no conflict of interest to disclose.
Figures
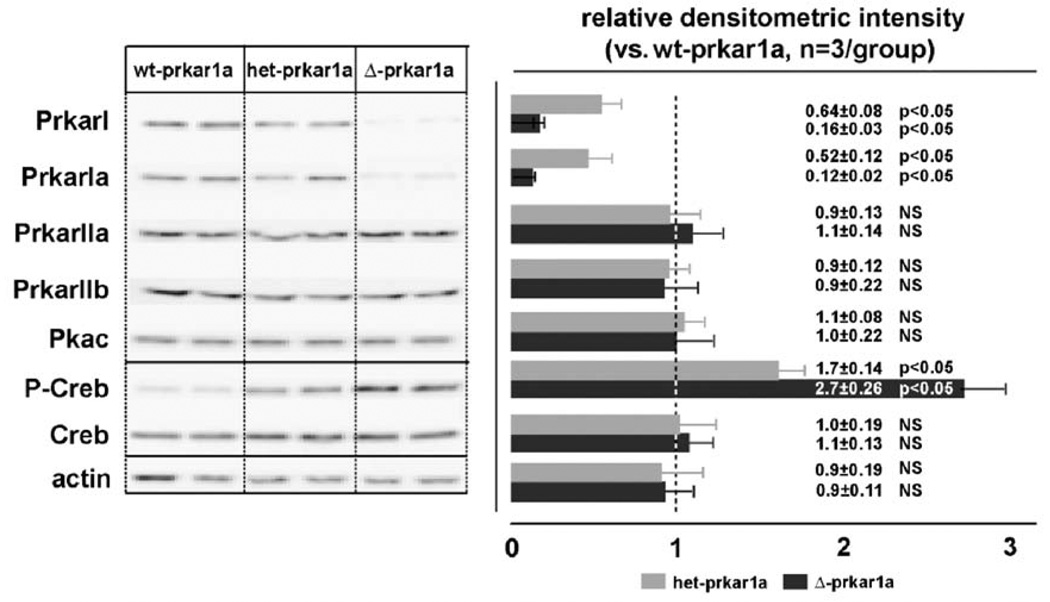
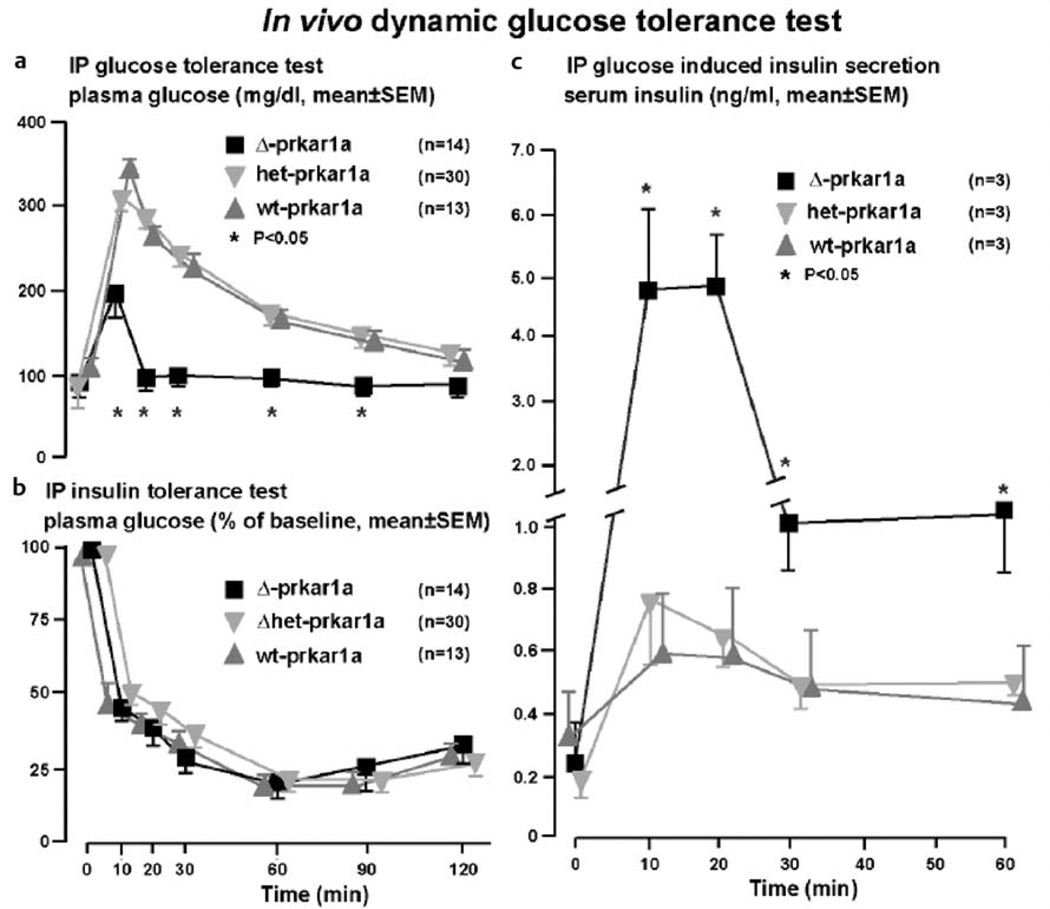
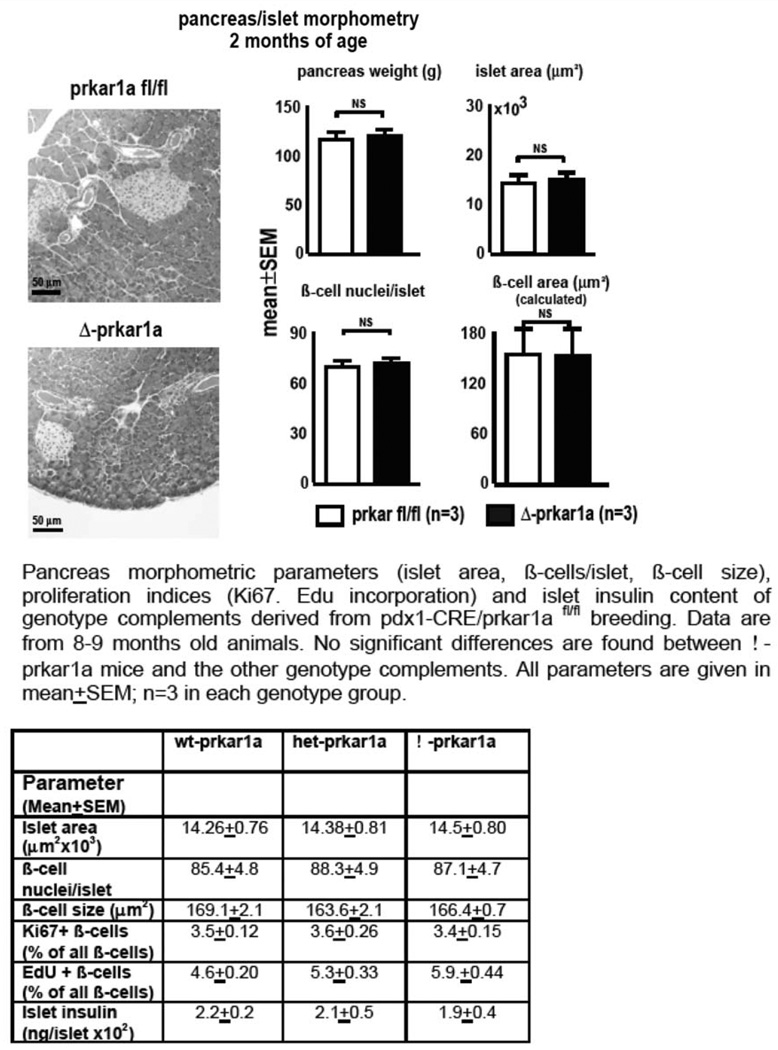
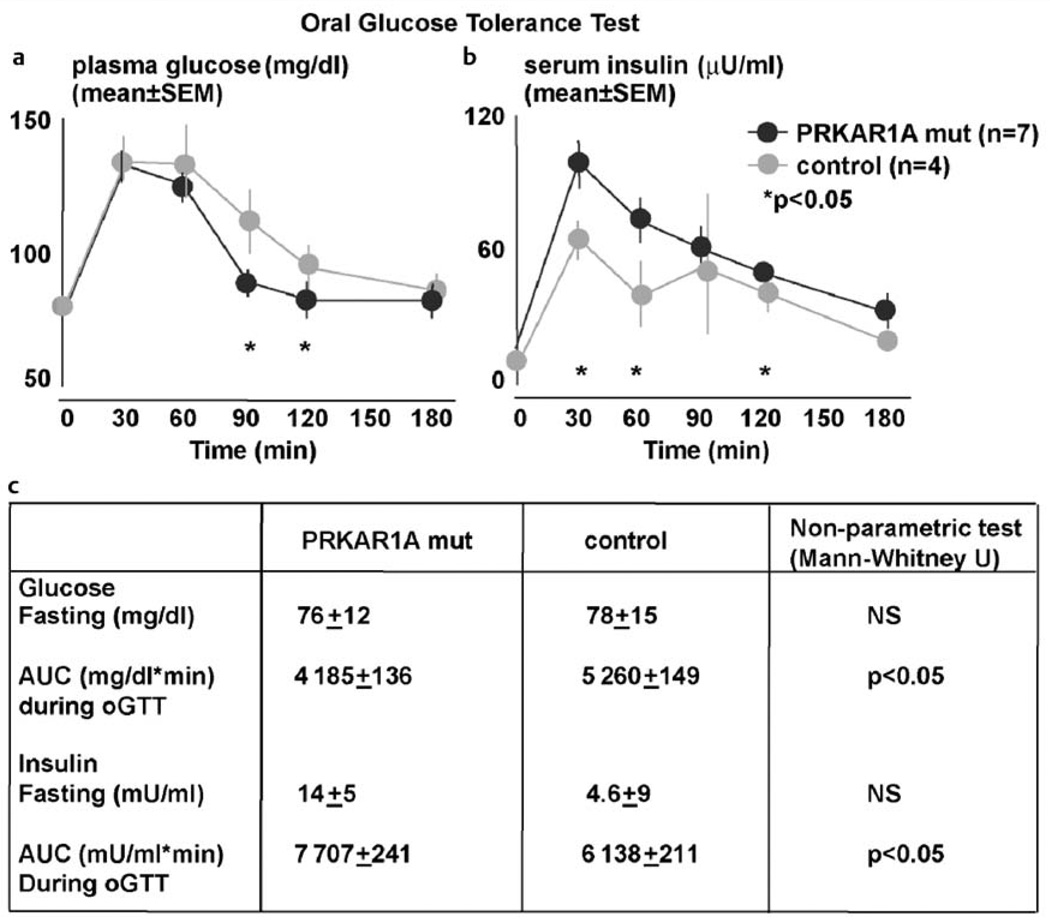
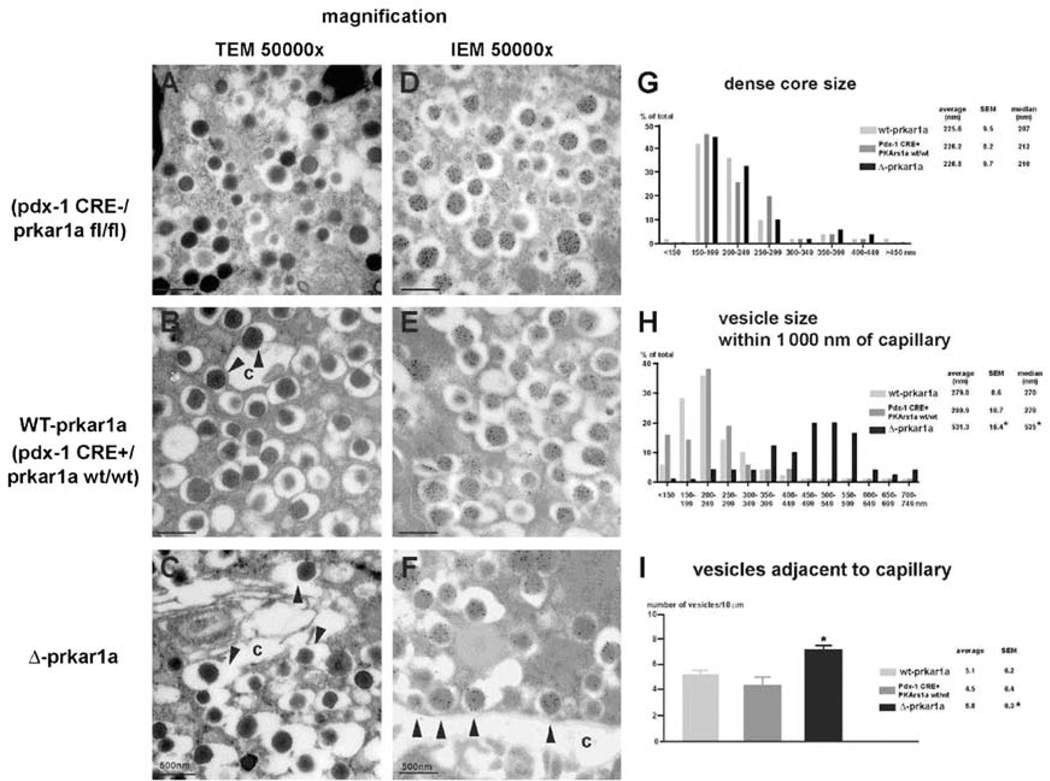
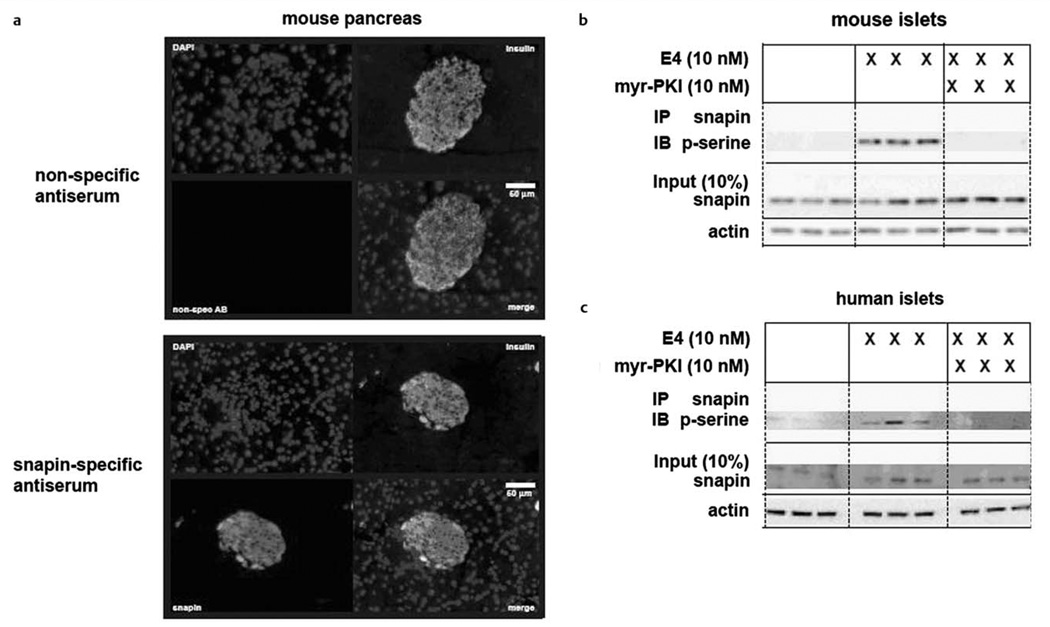

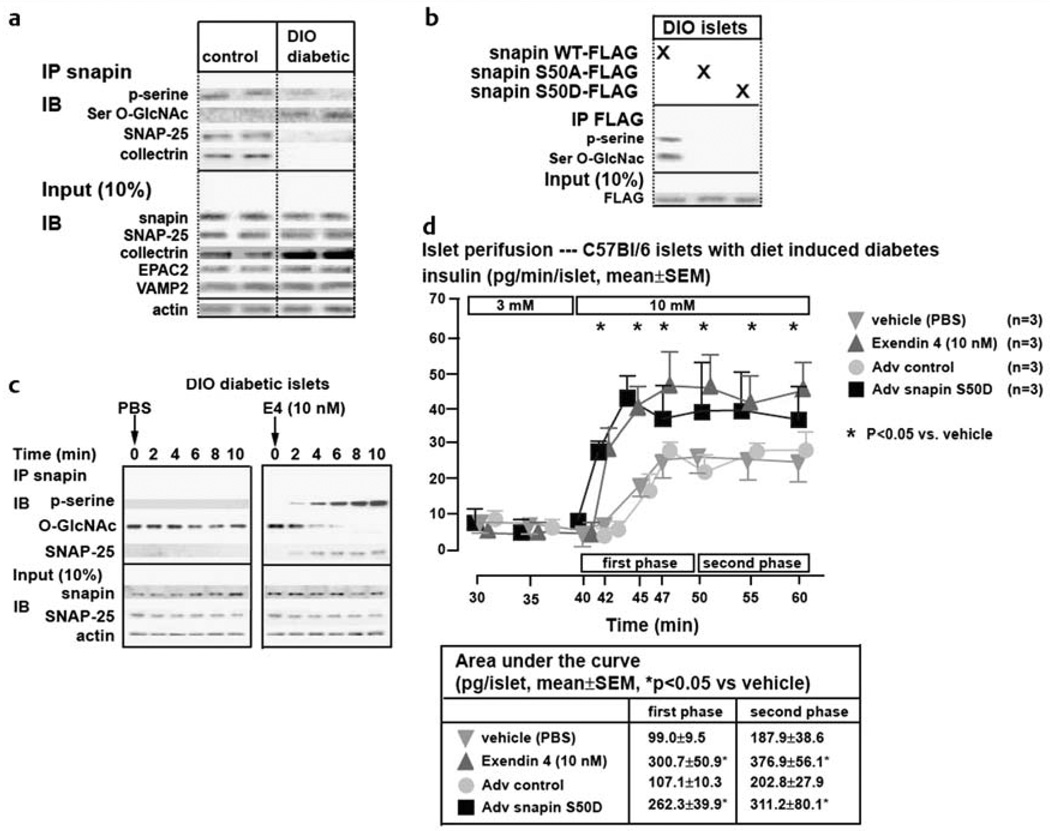
Similar articles
-
Snapin mediates incretin action and augments glucose-dependent insulin secretion.Cell Metab. 2011 Mar 2;13(3):308-19. doi: 10.1016/j.cmet.2011.02.002. Cell Metab. 2011. PMID: 21356520 Free PMC article.
-
Phospholipase C-ε links Epac2 activation to the potentiation of glucose-stimulated insulin secretion from mouse islets of Langerhans.Islets. 2011 May-Jun;3(3):121-8. doi: 10.4161/isl.3.3.15507. Epub 2011 May 1. Islets. 2011. PMID: 21478675 Free PMC article.
-
Glucotoxicity inhibits cAMP-protein kinase A-potentiated glucose-stimulated insulin secretion in pancreatic β-cells.J Diabetes. 2015 May;7(3):378-85. doi: 10.1111/1753-0407.12185. Epub 2014 Sep 6. J Diabetes. 2015. PMID: 24981285
-
Glucagon-like peptide 1-potentiated insulin secretion and proliferation of pancreatic β-cells.J Diabetes. 2014 Sep;6(5):394-402. doi: 10.1111/1753-0407.12161. Epub 2014 May 22. J Diabetes. 2014. PMID: 24725840 Review.
-
Anti-diabetic actions of glucagon-like peptide-1 on pancreatic beta-cells.Metabolism. 2014 Jan;63(1):9-19. doi: 10.1016/j.metabol.2013.09.010. Epub 2013 Oct 17. Metabolism. 2014. PMID: 24140094 Review.
Cited by
-
Cyclic AMP, protein kinase A, and phosphodiesterases: proceedings of an international workshop.Horm Metab Res. 2012 Sep;44(10):713-5. doi: 10.1055/s-0031-1301327. Epub 2012 Sep 5. Horm Metab Res. 2012. PMID: 22951901 Free PMC article.
-
Noncanonical Regulation of cAMP-Dependent Insulin Secretion and Its Implications in Type 2 Diabetes.Compr Physiol. 2023 Jun 26;13(3):5023-5049. doi: 10.1002/cphy.c220031. Compr Physiol. 2023. PMID: 37358504 Free PMC article.
-
Identification of Key Genes and Imbalanced SNAREs Assembly in the Comorbidity of Polycystic Ovary Syndrome and Depression.Genes (Basel). 2024 Apr 15;15(4):494. doi: 10.3390/genes15040494. Genes (Basel). 2024. PMID: 38674428 Free PMC article.
-
Significance of circulating microRNAs in diabetes mellitus type 2 and platelet reactivity: bioinformatic analysis and review.Cardiovasc Diabetol. 2019 Aug 30;18(1):113. doi: 10.1186/s12933-019-0918-x. Cardiovasc Diabetol. 2019. PMID: 31470851 Free PMC article. Review.
-
Restoration of Glucose-Stimulated Cdc42-Pak1 Activation and Insulin Secretion by a Selective Epac Activator in Type 2 Diabetic Human Islets.Diabetes. 2018 Oct;67(10):1999-2011. doi: 10.2337/db17-1174. Epub 2018 Jul 9. Diabetes. 2018. PMID: 29986926 Free PMC article.
References
-
- Nolan CJ, Damm P, Prentki M. Type 2 diabetes across generations: from pathophysiology to prevention and management. Lancet. 2011;378:169–181. - PubMed
-
- Del Prato S, Tiengo A. The importance of first-phase insulin secretion: implications for the therapy of type 2 diabetes mellitus. Diabetes Metab Res Rev. 2001;17:164–174. - PubMed
-
- Gerich JE. Is reduced first-phase insulin release the earliest detectable abnormality in individuals destined to develop type 2 diabetes? Diabetes. 2002;51(Suppl 1):S117–S121. - PubMed
-
- Lillioja S, Mott DM, Howard BV, Bennett PH, Yki-Jarvinen H, Frimond D, Nyomba BL, Zurlo F, Swinburn B, Bogardus C. Impaired glucose tolerance as a disorder of insulin action. Longitudinal and cross-sectional studies in Pima Indians. N Engl J Med. 1988;318:1217–1225. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical