

Bundling actin filaments from membranes: some novel players
- PMID: 22936939
- PMCID: PMC3426786
- DOI: 10.3389/fpls.2012.00188
Bundling actin filaments from membranes: some novel players
Abstract
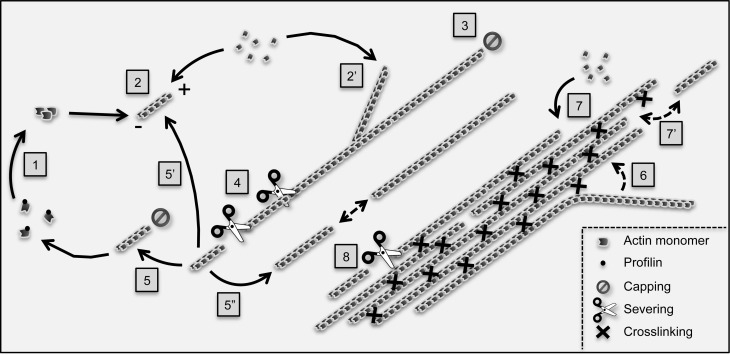
Progress in live-cell imaging of the cytoskeleton has significantly extended our knowledge about the organization and dynamics of actin filaments near the plasma membrane of plant cells. Noticeably, two populations of filamentous structures can be distinguished. On the one hand, fine actin filaments which exhibit an extremely dynamic behavior basically characterized by fast polymerization and prolific severing events, a process referred to as actin stochastic dynamics. On the other hand, thick actin bundles which are composed of several filaments and which are comparatively more stable although they constantly remodel as well. There is evidence that the actin cytoskeleton plays critical roles in trafficking and signaling at both the cell cortex and organelle periphery but the exact contribution of actin bundles remains unclear. A common view is that actin bundles provide the long-distance tracks used by myosin motors to deliver their cargo to growing regions and accordingly play a particularly important role in cell polarization. However, several studies support that actin bundles are more than simple passive highways and display multiple and dynamic roles in the regulation of many processes, such as cell elongation, polar auxin transport, stomatal and chloroplast movement, and defense against pathogens. The list of identified plant actin-bundling proteins is ever expanding, supporting that plant cells shape structurally and functionally different actin bundles. Here I review the most recently characterized actin-bundling proteins, with a particular focus on those potentially relevant to membrane trafficking and/or signaling.
Keywords: LIM proteins; SCAB1; THRUMIN1; V-ATPases; actin bundling; fimbrins; formins; villins.
Figures
Similar articles
-
Actin bundling in plants.Cell Motil Cytoskeleton. 2009 Nov;66(11):940-57. doi: 10.1002/cm.20389. Cell Motil Cytoskeleton. 2009. PMID: 19504571 Review.
-
Stochastic dynamics of actin filaments in guard cells regulating chloroplast localization during stomatal movement.Plant Cell Environ. 2011 Aug;34(8):1248-57. doi: 10.1111/j.1365-3040.2011.02325.x. Epub 2011 Apr 21. Plant Cell Environ. 2011. PMID: 21443604
-
THRUMIN1 is a light-regulated actin-bundling protein involved in chloroplast motility.Curr Biol. 2011 Jan 11;21(1):59-64. doi: 10.1016/j.cub.2010.11.059. Epub 2010 Dec 23. Curr Biol. 2011. PMID: 21185188
-
Microfilaments in cellular and developmental processes.Science. 1971 Jan 15;171(3967):135-43. doi: 10.1126/science.171.3967.135. Science. 1971. PMID: 5538822
-
Critical role of actin bundling in plant cell morphogenesis.Plant Signal Behav. 2010 May;5(5):484-8. doi: 10.4161/psb.10947. Epub 2010 Feb 1. Plant Signal Behav. 2010. PMID: 20118661 Free PMC article. Review.
Cited by
-
Automatic extraction of actin networks in plants.PLoS Comput Biol. 2023 Aug 30;19(8):e1011407. doi: 10.1371/journal.pcbi.1011407. eCollection 2023 Aug. PLoS Comput Biol. 2023. PMID: 37647341 Free PMC article.
-
TWISTED DWARF1 Mediates the Action of Auxin Transport Inhibitors on Actin Cytoskeleton Dynamics.Plant Cell. 2016 Apr;28(4):930-48. doi: 10.1105/tpc.15.00726. Epub 2016 Apr 6. Plant Cell. 2016. PMID: 27053424 Free PMC article.
-
The plasma membrane-associated Ca2+ -binding protein, PCaP1, is required for oligogalacturonide and flagellin-induced priming and immunity.Plant Cell Environ. 2021 Sep;44(9):3078-3093. doi: 10.1111/pce.14118. Epub 2021 Jun 30. Plant Cell Environ. 2021. PMID: 34050546 Free PMC article.
-
Polar Expansion Dynamics in the Plant Kingdom: A Diverse and Multifunctional Journey on the Path to Pollen Tubes.Plants (Basel). 2013 Mar 18;2(1):148-73. doi: 10.3390/plants2010148. Plants (Basel). 2013. PMID: 27137370 Free PMC article. Review.
-
The Pseudomonas syringae Type III Effector HopG1 Induces Actin Remodeling to Promote Symptom Development and Susceptibility during Infection.Plant Physiol. 2016 Jul;171(3):2239-55. doi: 10.1104/pp.16.01593. Epub 2016 May 23. Plant Physiol. 2016. PMID: 27217495 Free PMC article.
References
-
- Adams D. S., Robinson K. R., Fukumoto T., Yuan S., Albertson R. C., Yelick P., Kuo L., McSweeney M., Levin M. (2006). Early, H+-V-ATPase-dependent proton flux is necessary for consistent left-right patterning of non-mammalian vertebrates. Development 133, 1657–1671 10.1242/dev.02341 - DOI - PMC - PubMed
-
- Al Tanoury Z., Schaffner-Reckinger E., Halavatyi A., Hoffmann C., Moes M., Hadzic E., Catillon M., Yatskou M., Friederich E. (2010). Quantitative kinetic study of the actin-bundling protein L-plastin and of its impact on actin turn-over. PLoS ONE 5:e9210 10.1371/journal.pone.0009210 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources