

Identification of the HIV-1 NC binding interface in Alix Bro1 reveals a role for RNA
- PMID: 22896625
- PMCID: PMC3486338
- DOI: 10.1128/JVI.01260-12
Identification of the HIV-1 NC binding interface in Alix Bro1 reveals a role for RNA
Abstract
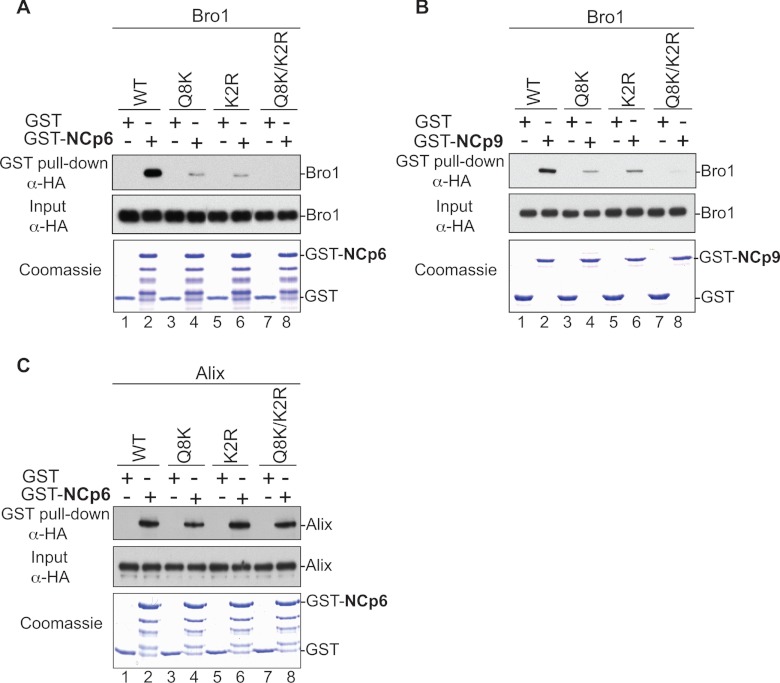
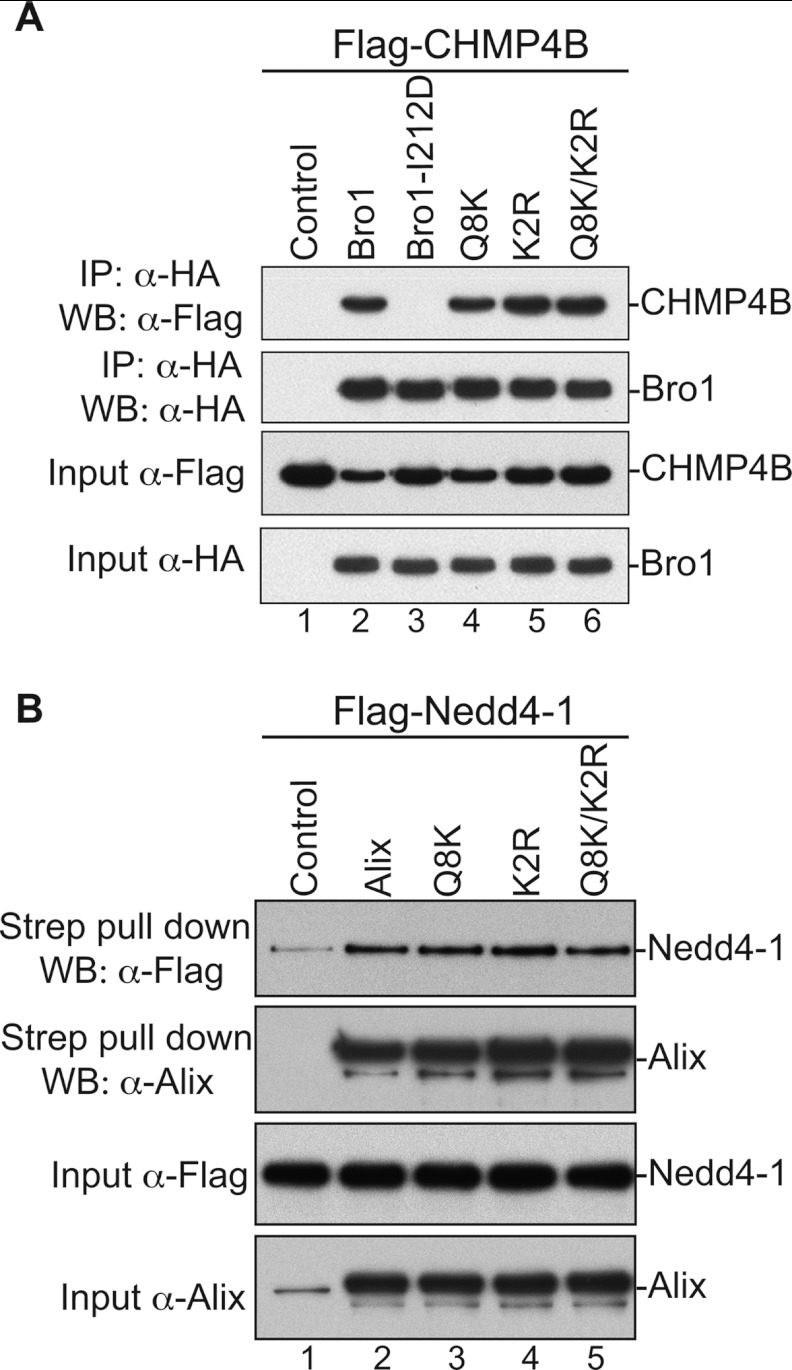
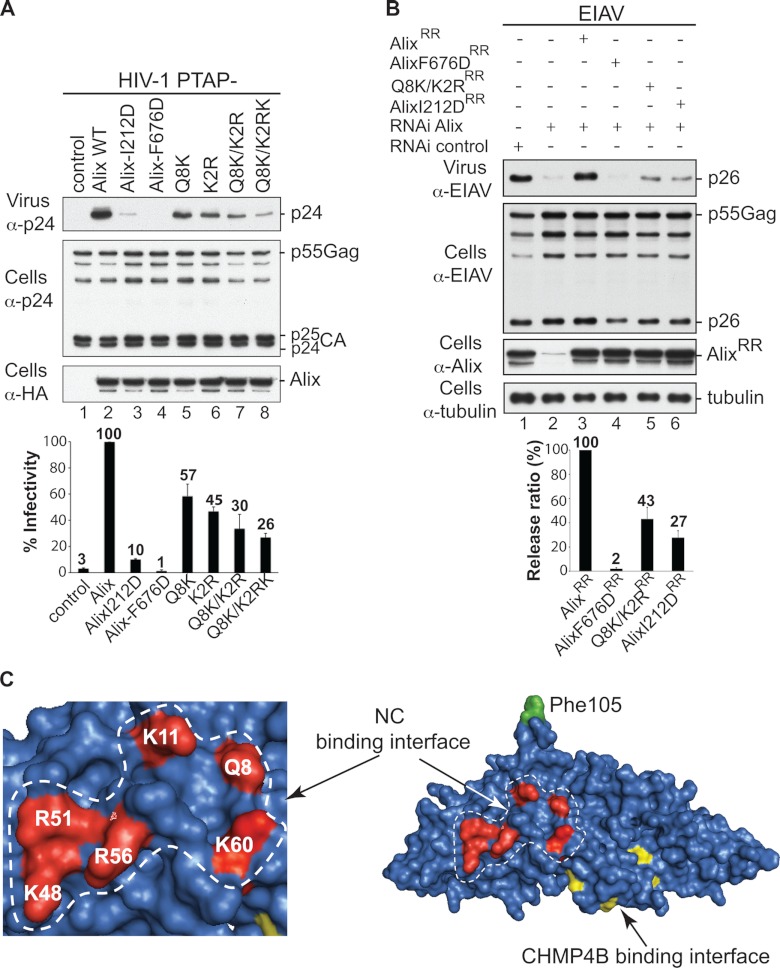
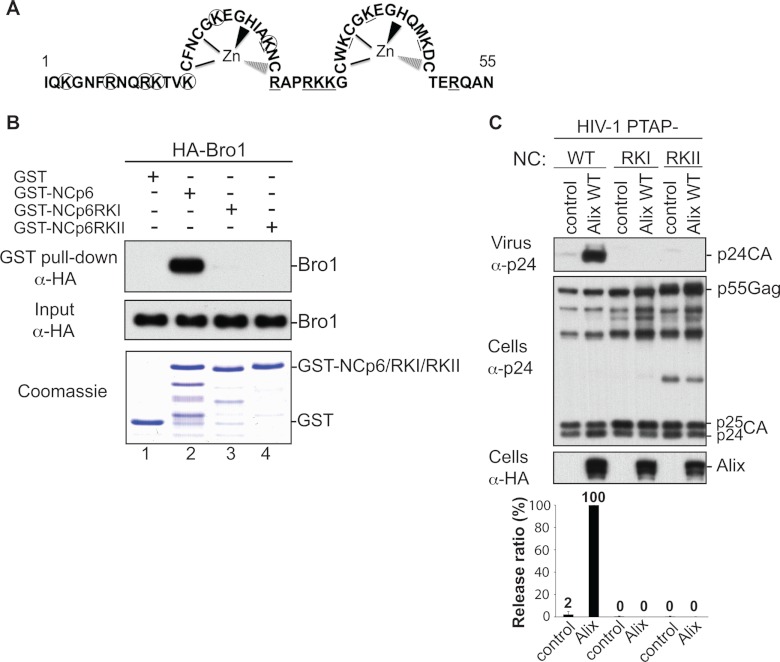
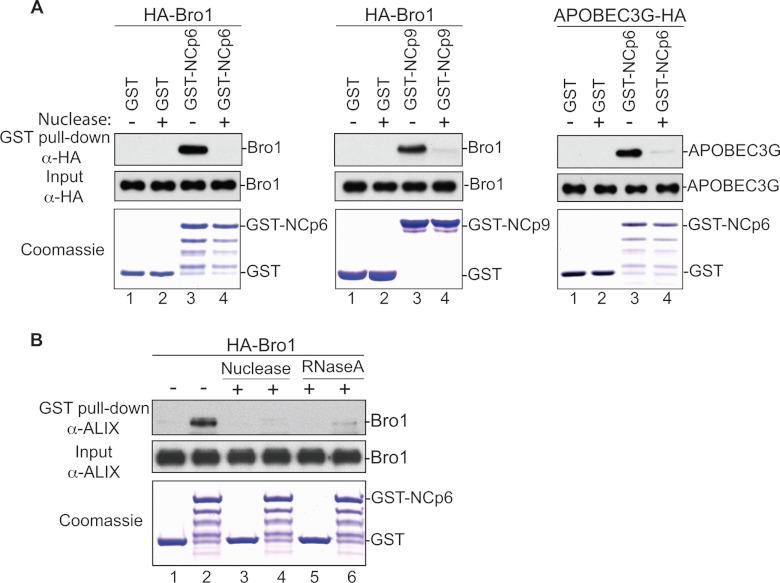
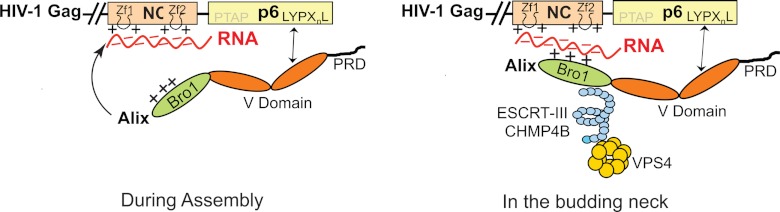
HIV-1 recruits members of ESCRT, the cell membrane fission machinery that promotes virus exit. HIV-1 Gag protein gains access to ESCRT directly by binding Alix, an ESCRT-associated protein that promotes budding. The Alix Bro1 and V domains bind Gag NC and p6 regions, respectively. Whereas V-p6 binding and function are well characterized, residues in Bro1 that interact with NC and their functional contribution to Alix-mediated HIV-1 budding are unknown. We mapped Bro1 residues that constitute the NC binding interface and found that they are critical for function. Intriguingly, residues involved in interactions on both sides of the Bro1-NC interface are positively charged, suggesting the involvement of a negatively charged cellular factor serving as a bridge. Nuclease treatment eliminated Bro1-NC interactions, revealing the involvement of RNA. These findings establish a direct role for NC in mediating interactions with ESCRT necessary for virus release and report the first evidence of RNA involvement in such recruitments.
Figures
Similar articles
-
Alix-Mediated Rescue of Feline Immunodeficiency Virus Budding Differs from That Observed with Human Immunodeficiency Virus.J Virol. 2020 May 18;94(11):e02019-19. doi: 10.1128/JVI.02019-19. Print 2020 May 18. J Virol. 2020. PMID: 32213612 Free PMC article.
-
The ESCRT-associated protein Alix recruits the ubiquitin ligase Nedd4-1 to facilitate HIV-1 release through the LYPXnL L domain motif.J Virol. 2010 Aug;84(16):8181-92. doi: 10.1128/JVI.00634-10. Epub 2010 Jun 2. J Virol. 2010. PMID: 20519395 Free PMC article.
-
The nucleocapsid region of HIV-1 Gag cooperates with the PTAP and LYPXnL late domains to recruit the cellular machinery necessary for viral budding.PLoS Pathog. 2009 Mar;5(3):e1000339. doi: 10.1371/journal.ppat.1000339. Epub 2009 Mar 13. PLoS Pathog. 2009. PMID: 19282983 Free PMC article.
-
Wrapping up the bad news: HIV assembly and release.Retrovirology. 2013 Jan 10;10:5. doi: 10.1186/1742-4690-10-5. Retrovirology. 2013. PMID: 23305486 Free PMC article. Review.
-
The role of cellular factors in promoting HIV budding.J Mol Biol. 2011 Jul 22;410(4):525-33. doi: 10.1016/j.jmb.2011.04.055. J Mol Biol. 2011. PMID: 21762798 Free PMC article. Review.
Cited by
-
RNA Binding Suppresses Tsg101 Recognition of Ub-Modified Gag and Facilitates Recruitment to the Plasma Membrane.Viruses. 2020 Apr 15;12(4):447. doi: 10.3390/v12040447. Viruses. 2020. PMID: 32326417 Free PMC article.
-
Viral membrane scission.Annu Rev Cell Dev Biol. 2013;29:551-69. doi: 10.1146/annurev-cellbio-101011-155838. Epub 2013 May 31. Annu Rev Cell Dev Biol. 2013. PMID: 24099087 Free PMC article. Review.
-
HIV Genome-Wide Protein Associations: a Review of 30 Years of Research.Microbiol Mol Biol Rev. 2016 Jun 29;80(3):679-731. doi: 10.1128/MMBR.00065-15. Print 2016 Sep. Microbiol Mol Biol Rev. 2016. PMID: 27357278 Free PMC article. Review.
-
Modulation of the HIV nucleocapsid dynamics finely tunes its RNA-binding properties during virion genesis.Nucleic Acids Res. 2018 Oct 12;46(18):9699-9710. doi: 10.1093/nar/gky612. Nucleic Acids Res. 2018. PMID: 29986076 Free PMC article.
-
Role of Alix in miRNA packaging during extracellular vesicle biogenesis.Int J Mol Med. 2016 Apr;37(4):958-66. doi: 10.3892/ijmm.2016.2488. Epub 2016 Feb 12. Int J Mol Med. 2016. PMID: 26935291 Free PMC article.
References
-
- Babst M, Katzmann DJ, Estepa-Sabal EJ, Meerloo T, Emr SD. 2002. Escrt-III: an endosome-associated heterooligomeric protein complex required for mvb sorting. Dev. Cell 3:271–282 - PubMed
-
- Babst M, Odorizzi G, Estepa EJ, Emr SD. 2000. Mammalian tumor susceptibility gene 101 (TSG101) and the yeast homologue, Vps23p, both function in late endosomal trafficking. Traffic 1:248–258 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
