

Purkinje cell dysfunction and delayed death in plasma membrane calcium ATPase 2-heterozygous mice
- PMID: 22789621
- PMCID: PMC3436994
- DOI: 10.1016/j.mcn.2012.07.001
Purkinje cell dysfunction and delayed death in plasma membrane calcium ATPase 2-heterozygous mice
Abstract
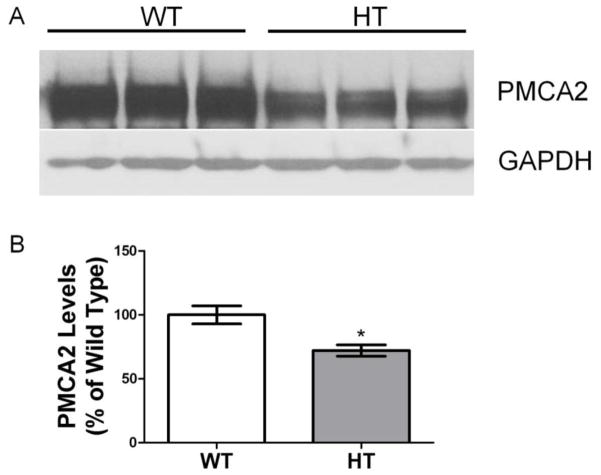
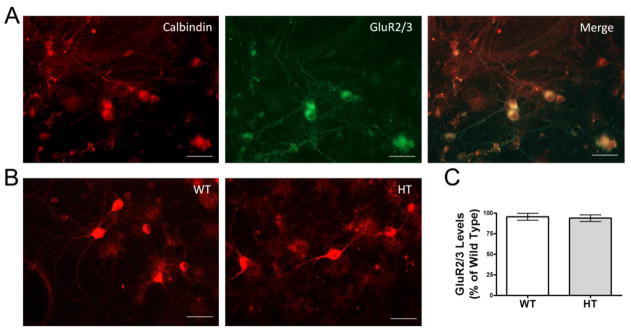
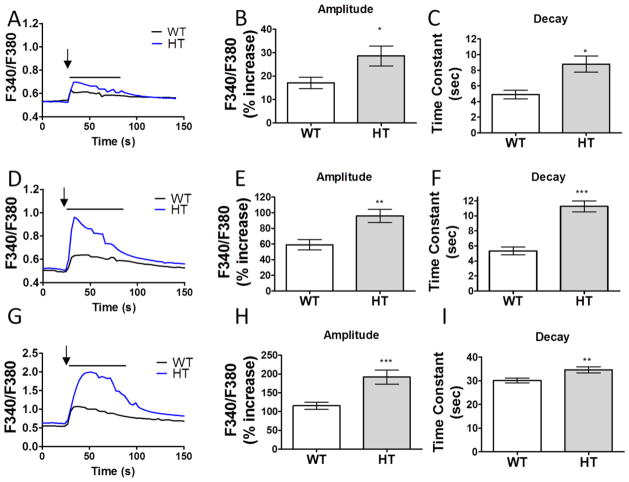
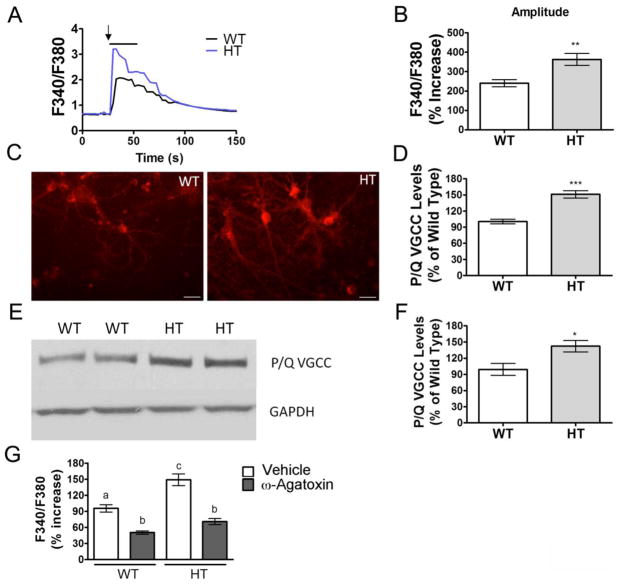
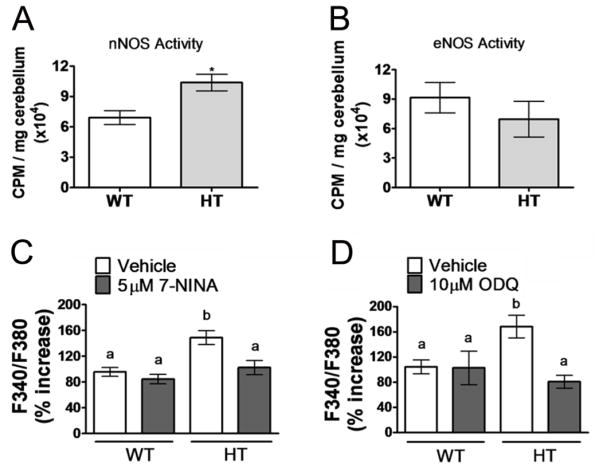
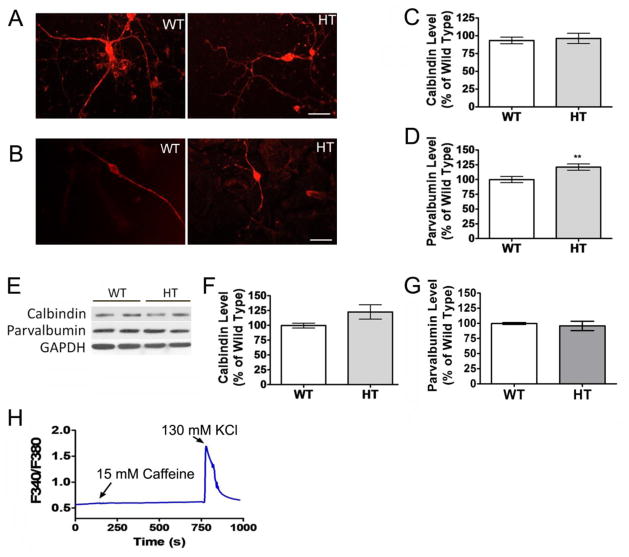
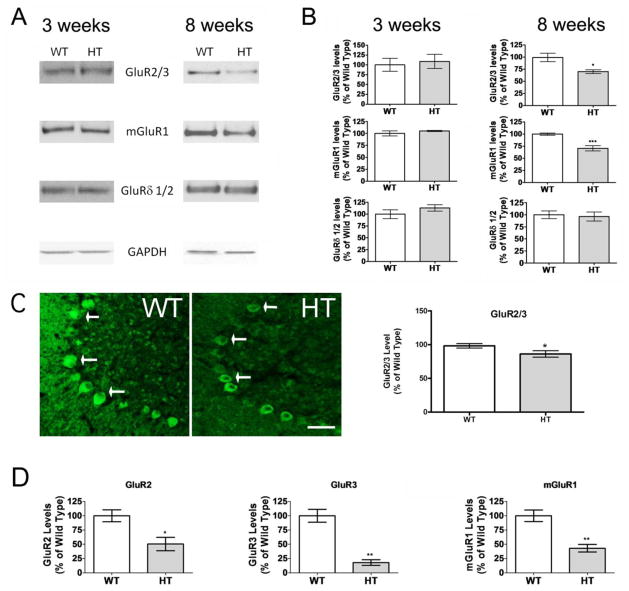
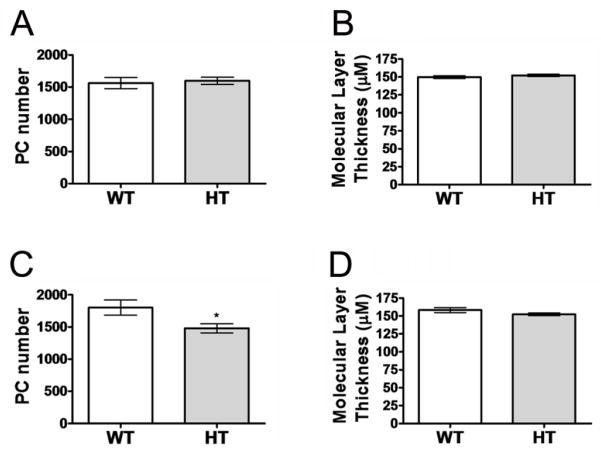
Purkinje cell (PC) dysfunction or death has been implicated in a number of disorders including ataxia, autism and multiple sclerosis. Plasma membrane calcium ATPase 2 (PMCA2), an important calcium (Ca(2+)) extrusion pump that interacts with synaptic signaling complexes, is most abundantly expressed in PCs compared to other neurons. Using the PMCA2 heterozygous mouse as a model, we investigated whether a reduction in PMCA2 levels affects PC function. We focused on Ca(2+) signaling and the expression of glutamate receptors which play a key role in PC function including synaptic plasticity. We found that the amplitude of depolarization and 2-amino-3-(5-methyl-3-oxo-1,2-oxazol-4-yl)propanoic acid receptor (AMPAR)-mediated Ca(2+) transients are significantly higher in cultured PMCA2(+/-) PCs than in PMCA2(+/+) PCs. This is due to increased Ca(2+) influx, since P/Q type voltage-gated Ca(2+) channel (VGCC) expression was more pronounced in PCs and cerebella of PMCA2(+/-) mice and VGCC blockade prevented the elevation in amplitude. Neuronal nitric oxide synthase (nNOS) activity was higher in PMCA2(+/-) cerebella and inhibition of nNOS or the soluble guanylate cyclase (sGC)-cyclic guanosine monophosphate (cGMP) pathway, which mediates nitric oxide (NO) signaling, reduced the amplitude of Ca(2+) transients in PMCA2(+/-) PCs, in vitro. In addition, there was an age-dependent decrease in metabotropic glutamate receptor 1 (mGluR1) and AMPA receptor subunit GluR2/3 transcript and protein levels at 8 weeks of age. These changes were followed by PC loss in the 20-week-old PMCA2(+/-) mice. Our studies highlight the importance of PMCA2 in Ca(2+) signaling, glutamate receptor expression and survival of Purkinje cells.
Copyright © 2012 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Molecular alterations in the cerebellum of the plasma membrane calcium ATPase 2 (PMCA2)-null mouse indicate abnormalities in Purkinje neurons.Mol Cell Neurosci. 2007 Feb;34(2):178-88. doi: 10.1016/j.mcn.2006.10.010. Epub 2006 Dec 5. Mol Cell Neurosci. 2007. PMID: 17150372 Free PMC article.
-
The plasma membrane Ca2+-ATPase2 (PMCA2) is involved in the regulation of Purkinje cell dendritic growth in cerebellar organotypic slice cultures.Neural Plast. 2013;2013:321685. doi: 10.1155/2013/321685. Epub 2013 Sep 29. Neural Plast. 2013. PMID: 24288624 Free PMC article.
-
Reduced expression of the Ca(2+) transporter protein PMCA2 slows Ca(2+) dynamics in mouse cerebellar Purkinje neurones and alters the precision of motor coordination.J Physiol. 2010 Mar 15;588(Pt 6):907-22. doi: 10.1113/jphysiol.2009.182196. Epub 2010 Jan 18. J Physiol. 2010. PMID: 20083513 Free PMC article.
-
Survival strategies for mouse cerebellar Purkinje neurons lacking PMCA2.Neurosci Lett. 2018 Jan 10;663:25-28. doi: 10.1016/j.neulet.2017.09.045. Neurosci Lett. 2018. PMID: 29452612 Review.
-
The plasma membrane calcium pump in the hearing process: physiology and pathology.Sci China Life Sci. 2011 Aug;54(8):686-90. doi: 10.1007/s11427-011-4200-z. Epub 2011 Jul 24. Sci China Life Sci. 2011. PMID: 21786191 Review.
Cited by
-
Evidence for a role of plasma membrane calcium pumps in neurodegenerative disease: Recent developments.Neurosci Lett. 2018 Jan 10;663:39-47. doi: 10.1016/j.neulet.2017.08.035. Epub 2017 Aug 19. Neurosci Lett. 2018. PMID: 28827127 Free PMC article. Review.
-
The evidence for association of ATP2B2 polymorphisms with autism in Chinese Han population.PLoS One. 2013 Apr 19;8(4):e61021. doi: 10.1371/journal.pone.0061021. Print 2013. PLoS One. 2013. PMID: 23620727 Free PMC article.
-
Alterations in Schizophrenia-Associated Genes Can Lead to Increased Power in Delta Oscillations.Cereb Cortex. 2019 Feb 1;29(2):875-891. doi: 10.1093/cercor/bhy291. Cereb Cortex. 2019. PMID: 30475994 Free PMC article.
-
Functional Effects of Schizophrenia-Linked Genetic Variants on Intrinsic Single-Neuron Excitability: A Modeling Study.Biol Psychiatry Cogn Neurosci Neuroimaging. 2016 Jan 1;1(1):49-59. doi: 10.1016/j.bpsc.2015.09.002. Biol Psychiatry Cogn Neurosci Neuroimaging. 2016. PMID: 26949748 Free PMC article.
-
Computational Modeling of Genetic Contributions to Excitability and Neural Coding in Layer V Pyramidal Cells: Applications to Schizophrenia Pathology.Front Comput Neurosci. 2019 Sep 26;13:66. doi: 10.3389/fncom.2019.00066. eCollection 2019. Front Comput Neurosci. 2019. PMID: 31616272 Free PMC article.
References
-
- Aiba A, Kano M, Chen C, Stanton ME, Fox GD, Herrup K, Zwingman TA, Tonegawa S. Deficient cerebellar long-term depression and impaired motor learning in mGluR1 mutant mice. Cell. 1994;79:377–388. - PubMed
-
- Bauman ML, Kemper TL. Neuroanatomic observations of the brain in autism: a review and future directions. Int J Dev Neurosci. 2005;23:183–187. - PubMed
-
- Burette A, Rockwood JM, Strehler EE, Weinberg RJ. Isoform-specific distribution of the plasma membrane Ca2+ ATPase in the rat brain. J Comp Neurol. 2003;467:464–476. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
