

Plectus - a stepping stone in embryonic cell lineage evolution of nematodes
- PMID: 22748136
- PMCID: PMC3464786
- DOI: 10.1186/2041-9139-3-13
Plectus - a stepping stone in embryonic cell lineage evolution of nematodes
Abstract
Background: Recent studies have challenged the widespread view that the pattern of embryogenesis found in Caenorhabditis elegans (clade 9) is characteristic of nematodes in general. To understand this still largely unexplored landscape of developmental events, we set out to examine more distantly related nematodes in detail for temporospatial differences in pattern formation and cell specification. Members of the genus Plectus (clade 6) seem to be suitable candidates to show variety, with certain idiosyncratic features during early development and the convenient availability of cultivatable species.
Methods: The study was conducted using 4-D lineage analysis, 3-D modeling of developing embryos and laser-induced ablation of individual blastomeres.
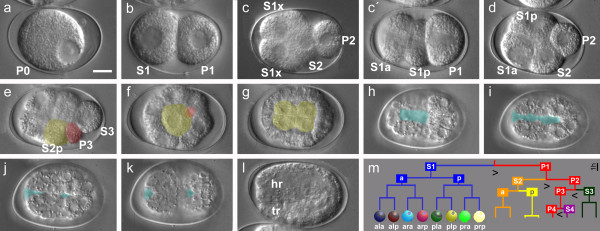
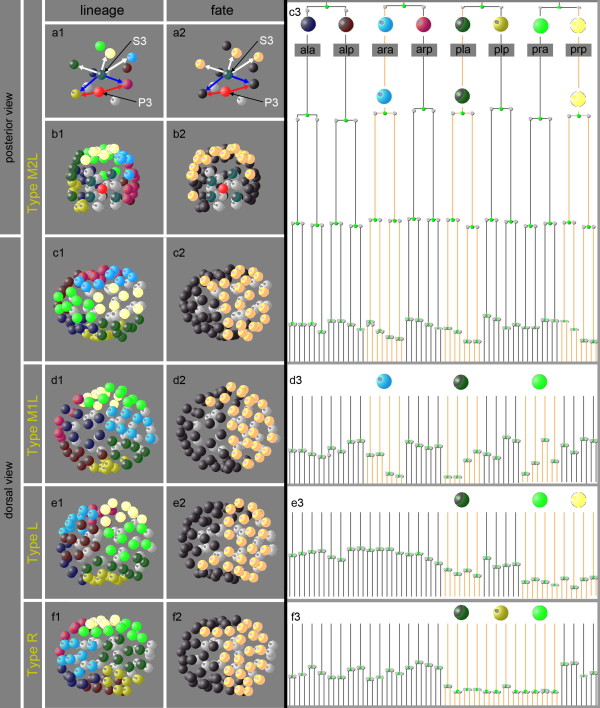
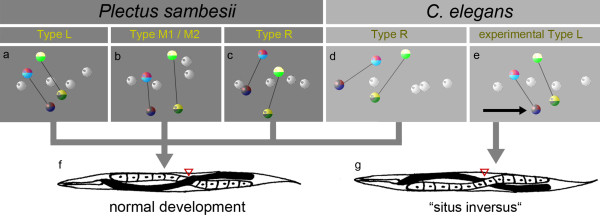
Results: Detailed cell lineage studies of several Plectus species reveal that pattern formation and cell fate assignment differ markedly from C. elegans. Descendants of the first somatic founder cell S1 (AB) - but not the progeny of other founder cells - demonstrate extremely variable spatial arrangements illustrating that here distinct early cell-cell interactions between invariant partners, as found in C. elegans, cannot take place. Different from C. elegans, in Plectus alternative positional variations among early S1 blastomeres resulting in a 'situs inversus' pattern, nevertheless give rise to adults with normal left-right asymmetries. In addition, laser ablations of early blastomeres uncover inductions between variable cell partners.
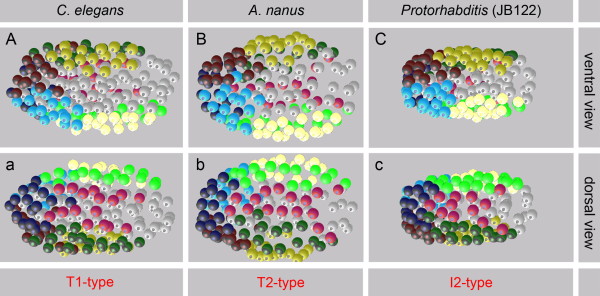
Conclusions: Our results suggest that embryonic cell specification in Plectus is not correlated with cell lineage but with position. With this peculiarity, Plectus appears to occupy an intermediate position between basal nematodes displaying a variable early development and the C. elegans-like invariant pattern. We suggest that indeterminate pattern formation associated with late, position-dependent fate assignment represents a plesiomorphic character among nematodes predominant in certain basal clades but lost in derived clades. Thus, the behavior of S1 cells in Plectus can be considered an evolutionary relict in a transition phase between two different developmental strategies.
Figures





Similar articles
-
Evolution of embryonic development in nematodes.Evodevo. 2011 Sep 20;2(1):18. doi: 10.1186/2041-9139-2-18. Evodevo. 2011. PMID: 21929824 Free PMC article.
-
Duels without obvious sense: counteracting inductions involved in body wall muscle development in the Caenorhabditis elegans embryo.Development. 1995 Jul;121(7):2219-32. doi: 10.1242/dev.121.7.2219. Development. 1995. PMID: 7635065
-
Specification of gut cell fate differs significantly between the nematodes Acrobeloides nanus and caenorhabditis elegans.Dev Biol. 1998 Dec 1;204(1):3-14. doi: 10.1006/dbio.1998.9054. Dev Biol. 1998. PMID: 9851839
-
Generation of cell diversity during early embryogenesis in the nematode Caenorhabditis elegans.Int Rev Cytol. 1989;114:81-123. doi: 10.1016/s0074-7696(08)60859-1. Int Rev Cytol. 1989. PMID: 2661459 Review.
-
[Specification of cell destiny in early Caenorhabditis elegans embryo].Naturwissenschaften. 1997 Feb;84(2):55-64. doi: 10.1007/s001140050349. Naturwissenschaften. 1997. PMID: 9121590 Review. German.
Cited by
-
The genome of Romanomermis culicivorax: revealing fundamental changes in the core developmental genetic toolkit in Nematoda.BMC Genomics. 2013 Dec 27;14:923. doi: 10.1186/1471-2164-14-923. BMC Genomics. 2013. PMID: 24373391 Free PMC article.
-
From "the Worm" to "the Worms" and Back Again: The Evolutionary Developmental Biology of Nematodes.Genetics. 2018 Oct;210(2):397-433. doi: 10.1534/genetics.118.300243. Genetics. 2018. PMID: 30287515 Free PMC article. Review.
-
Automated cell lineage reconstruction using label-free 4D microscopy.Genetics. 2024 Oct 7;228(2):iyae135. doi: 10.1093/genetics/iyae135. Genetics. 2024. PMID: 39139100 Free PMC article.
-
Laser-mediated cell ablation during post-implantation mouse development.Dev Dyn. 2013 Oct;242(10):1202-9. doi: 10.1002/dvdy.24017. Epub 2013 Sep 2. Dev Dyn. 2013. PMID: 23873840 Free PMC article.
-
Modulating cell proliferation by asymmetric division: A conserved pattern in the early embryogenesis of nematode species.MicroPubl Biol. 2024 Mar 4;2024:10.17912/micropub.biology.001006. doi: 10.17912/micropub.biology.001006. eCollection 2024. MicroPubl Biol. 2024. PMID: 38505394 Free PMC article.
References
-
- Blaxter M. In: Nematodes (Nematoda) Hedges SB, Kumar S, editor. Oxford University Press, Oxford; 2009. pp. 247–250.
-
- Poinar GO. The evolutionary history of nematodes: as revealed in stone, amber and mummies. Brill, Leiden; 2011.
-
- Poinar G, Kerp H, Hass H. Palaeonema phyticum gen. n., sp n. (Nematoda: Palaeonematidae fam. n.), a Devonian nematode associated with early land plants. Nematology. 2008;10:9–14.
-
- Meldal BH, Debenham NJ, De Ley P, De Ley IT, Vanfleteren JR, Vierstraete AR, Bert W, Borgonie G, Moens T, Tyler PA, Austen MC, Blaxter ML, Rogers AD, Lambshead PJ. An improved molecular phylogeny of the Nematoda with special emphasis on marine taxa. Mol Phylogenet Evol. 2007;42:622–636. - PubMed
LinkOut - more resources
Full Text Sources
