

E2F mediates enhanced alternative polyadenylation in proliferation
- PMID: 22747694
- PMCID: PMC3491381
- DOI: 10.1186/gb-2012-13-7-r59
E2F mediates enhanced alternative polyadenylation in proliferation
Abstract
Background: The majority of mammalian genes contain multiple poly(A) sites in their 3' UTRs. Alternative cleavage and polyadenylation are emerging as an important layer of gene regulation as they generate transcript isoforms that differ in their 3' UTRs, thereby modulating genes' response to 3' UTR-mediated regulation. Enhanced cleavage at 3' UTR proximal poly(A) sites resulting in global 3' UTR shortening was recently linked to proliferation and cancer. However, mechanisms that regulate this enhanced alternative polyadenylation are unknown.
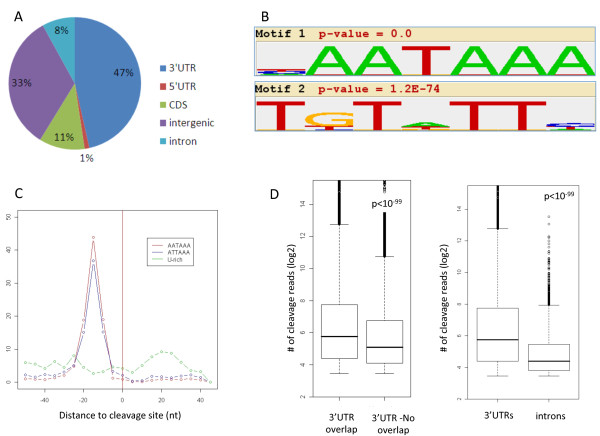
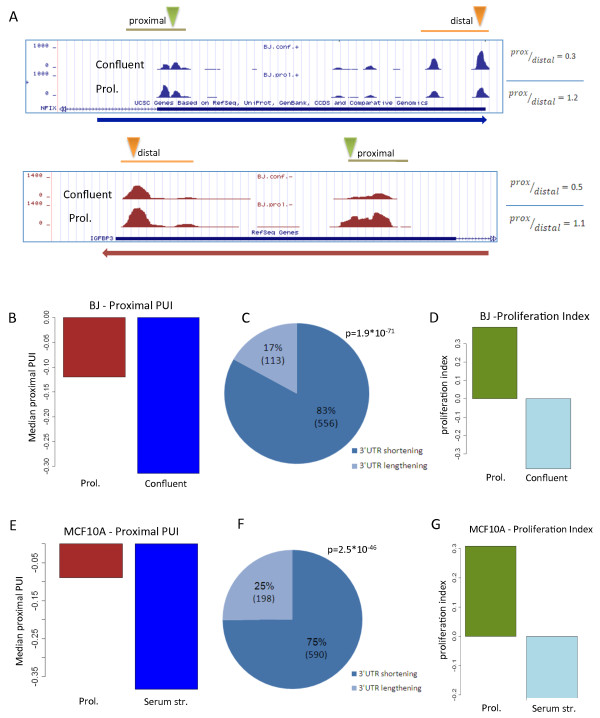
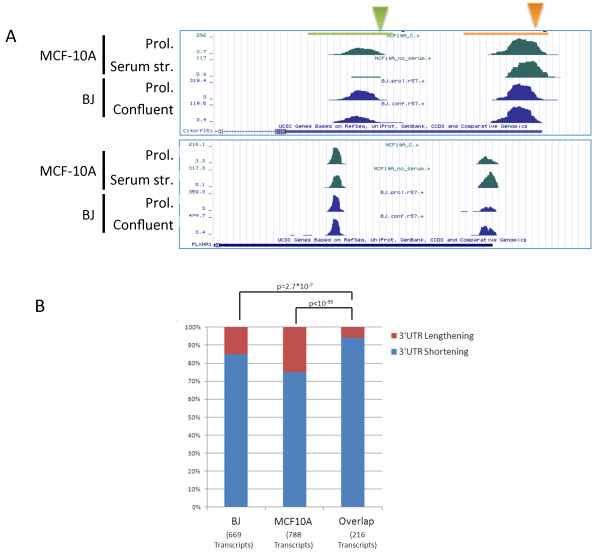
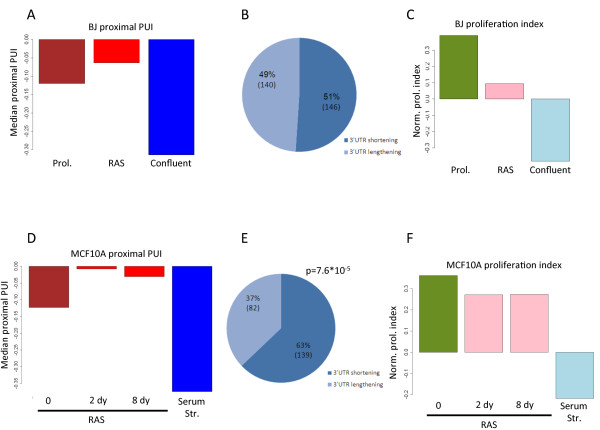
Results: Here, we explored, on a transcriptome-wide scale, alternative polyadenylation events associated with cellular proliferation and neoplastic transformation. We applied a deep-sequencing technique for identification and quantification of poly(A) sites to two human cellular models, each examined under proliferative, arrested and transformed states. In both cell systems we observed global 3' UTR shortening associated with proliferation, a link that was markedly stronger than the association with transformation. Furthermore, we found that proliferation is also associated with enhanced cleavage at intronic poly(A) sites. Last, we found that the expression level of the set of genes that encode for 3'-end processing proteins is globally elevated in proliferation, and that E2F transcription factors contribute to this regulation.
Conclusions: Our results comprehensively identify alternative polyadenylation events associated with cellular proliferation and transformation, and demonstrate that the enhanced alternative polyadenylation in proliferative conditions results not only in global 3' UTR shortening but also in enhanced premature cleavage in introns. Our results also indicate that E2F-mediated co-transcriptional regulation of 3'-end processing genes is one of the mechanisms that links enhanced alternative polyadenylation to proliferation.
Figures


Similar articles
-
Formation, regulation and evolution of Caenorhabditis elegans 3'UTRs.Nature. 2011 Jan 6;469(7328):97-101. doi: 10.1038/nature09616. Epub 2010 Nov 17. Nature. 2011. PMID: 21085120 Free PMC article.
-
CPEB1 coordinates alternative 3'-UTR formation with translational regulation.Nature. 2013 Mar 7;495(7439):121-5. doi: 10.1038/nature11901. Epub 2013 Feb 24. Nature. 2013. PMID: 23434754
-
Ubiquitously transcribed genes use alternative polyadenylation to achieve tissue-specific expression.Genes Dev. 2013 Nov 1;27(21):2380-96. doi: 10.1101/gad.229328.113. Epub 2013 Oct 21. Genes Dev. 2013. PMID: 24145798 Free PMC article.
-
Implications of polyadenylation in health and disease.Nucleus. 2014;5(6):508-19. doi: 10.4161/nucl.36360. Epub 2014 Oct 31. Nucleus. 2014. PMID: 25484187 Free PMC article. Review.
-
The Detection and Bioinformatic Analysis of Alternative 3' UTR Isoforms as Potential Cancer Biomarkers.Int J Mol Sci. 2021 May 18;22(10):5322. doi: 10.3390/ijms22105322. Int J Mol Sci. 2021. PMID: 34070203 Free PMC article. Review.
Cited by
-
Alternative cleavage and polyadenylation: the long and short of it.Trends Biochem Sci. 2013 Jun;38(6):312-20. doi: 10.1016/j.tibs.2013.03.005. Epub 2013 Apr 27. Trends Biochem Sci. 2013. PMID: 23632313 Free PMC article. Review.
-
Alternative polyadenylation regulation in cardiac development and cardiovascular disease.Cardiovasc Res. 2023 Jun 13;119(6):1324-1335. doi: 10.1093/cvr/cvad014. Cardiovasc Res. 2023. PMID: 36657944 Free PMC article. Review.
-
Sequence determinants as key regulators in gene expression of T cells.Immunol Rev. 2021 Nov;304(1):10-29. doi: 10.1111/imr.13021. Epub 2021 Sep 5. Immunol Rev. 2021. PMID: 34486113 Free PMC article. Review.
-
Dynamic alternative polyadenylation during iPSC differentiation into cardiomyocytes.Comput Struct Biotechnol J. 2022 Oct 25;20:5859-5869. doi: 10.1016/j.csbj.2022.10.025. eCollection 2022. Comput Struct Biotechnol J. 2022. PMID: 36382196 Free PMC article.
-
The role of alternative polyadenylation in breast cancer.Front Genet. 2024 Jun 13;15:1377275. doi: 10.3389/fgene.2024.1377275. eCollection 2024. Front Genet. 2024. PMID: 38939531 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
