

Evolutionary dynamics of RNA-like replicator systems: A bioinformatic approach to the origin of life
- PMID: 22727399
- PMCID: PMC3466355
- DOI: 10.1016/j.plrev.2012.06.001
Evolutionary dynamics of RNA-like replicator systems: A bioinformatic approach to the origin of life
Abstract
We review computational studies on prebiotic evolution, focusing on informatic processes in RNA-like replicator systems. In particular, we consider the following processes: the maintenance of information by replicators with and without interactions, the acquisition of information by replicators having a complex genotype-phenotype map, the generation of information by replicators having a complex genotype-phenotype-interaction map, and the storage of information by replicators serving as dedicated templates. Focusing on these informatic aspects, we review studies on quasi-species, error threshold, RNA-folding genotype-phenotype map, hypercycle, multilevel selection (including spatial self-organization, classical group selection, and compartmentalization), and the origin of DNA-like replicators. In conclusion, we pose a future question for theoretical studies on the origin of life.
Published by Elsevier B.V.
Figures
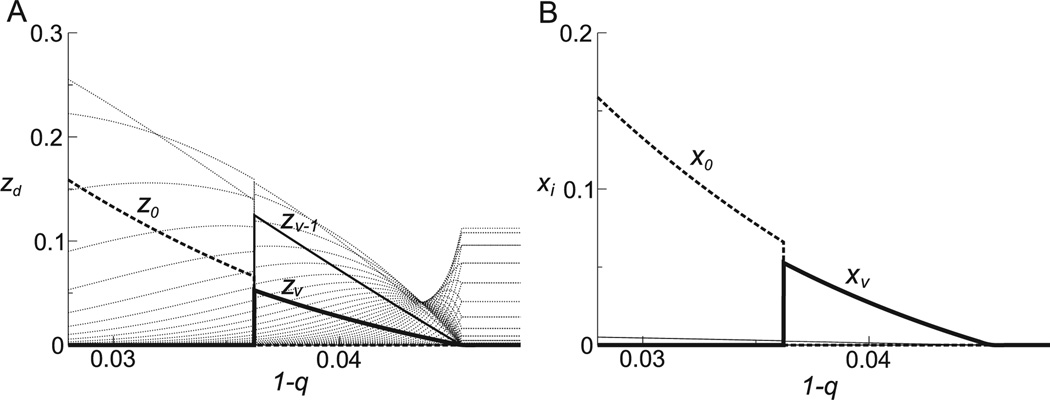











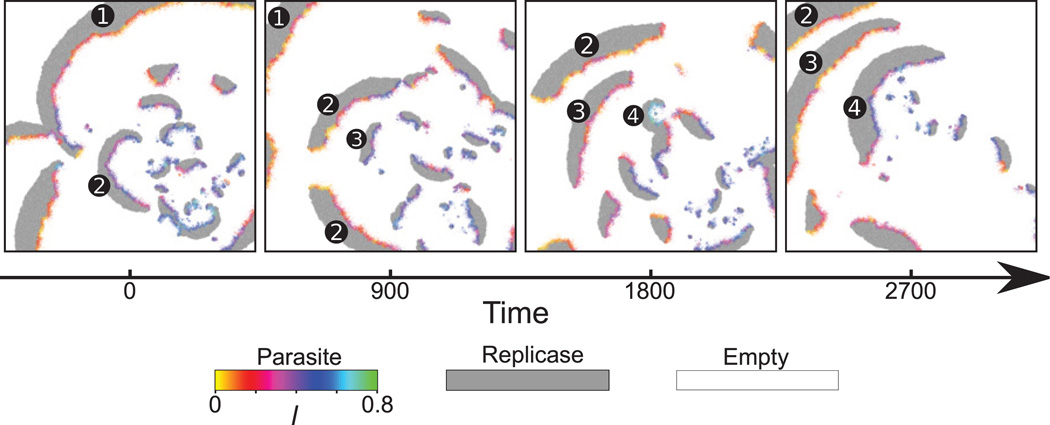









Comment in
-
Neutral networks and chemical function in RNA: Comment on "Evolutionary dynamics of RNA-like replicator systems: A bioinformatic approach to the origin of life".Phys Life Rev. 2012 Sep;9(3):277-8; discussion 279-84. doi: 10.1016/j.plrev.2012.06.003. Epub 2012 Jun 7. Phys Life Rev. 2012. PMID: 22698460 No abstract available.
-
Comment on "Evolutionary dynamics of RNA-like replicator systems: A bioinformatic approach to the origin of life" by Nobuto Takeuchi and Paulien Hogeweg.Phys Life Rev. 2012 Sep;9(3):268-9; discussion 279-84. doi: 10.1016/j.plrev.2012.06.006. Epub 2012 Jun 7. Phys Life Rev. 2012. PMID: 22698461 No abstract available.
-
How robust are the models of the origin of life?: Commentary on "Evolutionary dynamics of RNA-like replicator systems: A bioinformatic approach to the origin of life".Phys Life Rev. 2012 Sep;9(3):264-5; discussion 279-84. doi: 10.1016/j.plrev.2012.06.004. Epub 2012 Jun 7. Phys Life Rev. 2012. PMID: 22704231 No abstract available.
-
Survival of the pre-fittest: Commentary to N. Takeuchi and P. Hogeweg review article "Evolutionary dynamics of RNA-like replicator systems: A bioinformatic approach to the origin of life".Phys Life Rev. 2012 Sep;9(3):272-3; discussion 279-84. doi: 10.1016/j.plrev.2012.06.007. Epub 2012 Jun 8. Phys Life Rev. 2012. PMID: 22704836 No abstract available.
-
Comments on "Evolutionary dynamics of RNA-like replicator systems".Phys Life Rev. 2012 Sep;9(3):270-1; discussion 279-84. doi: 10.1016/j.plrev.2012.06.010. Epub 2012 Jun 8. Phys Life Rev. 2012. PMID: 22704837 No abstract available.
-
How universal is Darwin's principle?Phys Life Rev. 2012 Dec;9(4):460-1. doi: 10.1016/j.plrev.2012.06.009. Epub 2012 Jun 8. Phys Life Rev. 2012. PMID: 22727400 No abstract available.
-
Survival of the fittest or the flattest?: Commentary on "Evolutionary dynamics of RNA-like replicator systems: A bioinformatic approach to the origin of life" by Nobuto Takeuchi and Paulien Hogeweg.Phys Life Rev. 2012 Sep;9(3):266-7; discussion 279-84. doi: 10.1016/j.plrev.2012.06.008. Epub 2012 Jun 8. Phys Life Rev. 2012. PMID: 22727816 No abstract available.
-
The theoretical underpinnings of primordial RNA replication: Comment on "Evolutionary dynamics of RNA-like replicator systems: A bioinformatic approach to the origin of life" by Takeuchi and Hogeweg.Phys Life Rev. 2012 Sep;9(3):274-6; discussion 279-84. doi: 10.1016/j.plrev.2012.06.011. Epub 2012 Jun 21. Phys Life Rev. 2012. PMID: 22738813 No abstract available.
Similar articles
-
Evolution of complexity in RNA-like replicator systems.Biol Direct. 2008 Mar 27;3:11. doi: 10.1186/1745-6150-3-11. Biol Direct. 2008. PMID: 18371199 Free PMC article.
-
The theoretical underpinnings of primordial RNA replication: Comment on "Evolutionary dynamics of RNA-like replicator systems: A bioinformatic approach to the origin of life" by Takeuchi and Hogeweg.Phys Life Rev. 2012 Sep;9(3):274-6; discussion 279-84. doi: 10.1016/j.plrev.2012.06.011. Epub 2012 Jun 21. Phys Life Rev. 2012. PMID: 22738813 No abstract available.
-
Multilevel selection in models of prebiotic evolution II: a direct comparison of compartmentalization and spatial self-organization.PLoS Comput Biol. 2009 Oct;5(10):e1000542. doi: 10.1371/journal.pcbi.1000542. Epub 2009 Oct 16. PLoS Comput Biol. 2009. PMID: 19834556 Free PMC article.
-
Multilevel selection in models of prebiotic evolution: compartments and spatial self-organization.Orig Life Evol Biosph. 2003 Oct;33(4-5):375-403. doi: 10.1023/a:1025754907141. Orig Life Evol Biosph. 2003. PMID: 14604183 Review.
-
Evolution of RNA-Based Networks.Curr Top Microbiol Immunol. 2016;392:43-59. doi: 10.1007/82_2015_470. Curr Top Microbiol Immunol. 2016. PMID: 26336964 Review.
Cited by
-
Evolutionary transition from a single RNA replicator to a multiple replicator network.Nat Commun. 2022 Mar 18;13(1):1460. doi: 10.1038/s41467-022-29113-x. Nat Commun. 2022. PMID: 35304447 Free PMC article.
-
Virus world as an evolutionary network of viruses and capsidless selfish elements.Microbiol Mol Biol Rev. 2014 Jun;78(2):278-303. doi: 10.1128/MMBR.00049-13. Microbiol Mol Biol Rev. 2014. PMID: 24847023 Free PMC article. Review.
-
Evolution of microbes and viruses: a paradigm shift in evolutionary biology?Front Cell Infect Microbiol. 2012 Sep 13;2:119. doi: 10.3389/fcimb.2012.00119. eCollection 2012. Front Cell Infect Microbiol. 2012. PMID: 22993722 Free PMC article. Review.
-
Co-operation between Polymerases and Nucleotide Synthetases in the RNA World.PLoS Comput Biol. 2016 Nov 7;12(11):e1005161. doi: 10.1371/journal.pcbi.1005161. eCollection 2016 Nov. PLoS Comput Biol. 2016. PMID: 27820829 Free PMC article.
-
Survival of Self-Replicating Molecules under Transient Compartmentalization with Natural Selection.Life (Basel). 2019 Oct 3;9(4):78. doi: 10.3390/life9040078. Life (Basel). 2019. PMID: 31623412 Free PMC article.
References
-
- Phillips R, Kondev J, Theriot J. Physical Biology of the Cell. New York, NY, USA: Garland Science; 2009.
-
- Crick FHC. On protein synthesis. Symposia of the Society for Experimental Biology. 1958;12:138–163. - PubMed
-
- Crick F. Central dogma of molecular biology. Nature. 1970;227:561–563. - PubMed
-
- Moore PB, Steiz TA. The role of RNA in the synthesis of proteins. In: Gesteland RF, Cech TR, Atkins JF, editors. The RNA World. 3rd Edition. Cold Spring Harbor, NY, USA: Cold Spring Harbor Laboratory Press; 2005. pp. 257–285.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
