

The C-terminal cysteine annulus participates in auto-chaperone function for Salmonella phage P22 tailspike folding and assembly
- PMID: 22666655
- PMCID: PMC3357383
- DOI: 10.4161/bact.19775
The C-terminal cysteine annulus participates in auto-chaperone function for Salmonella phage P22 tailspike folding and assembly
Abstract
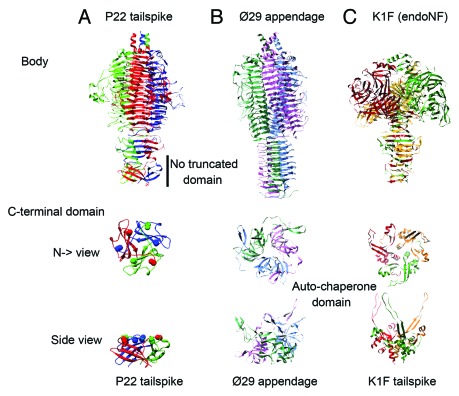
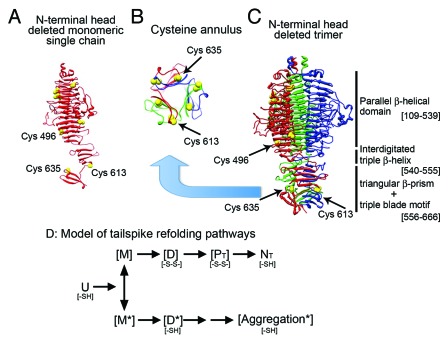
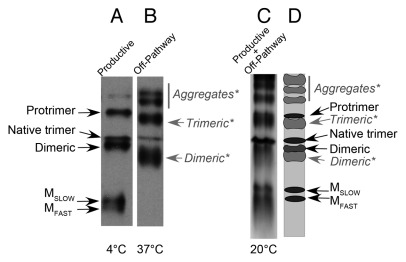
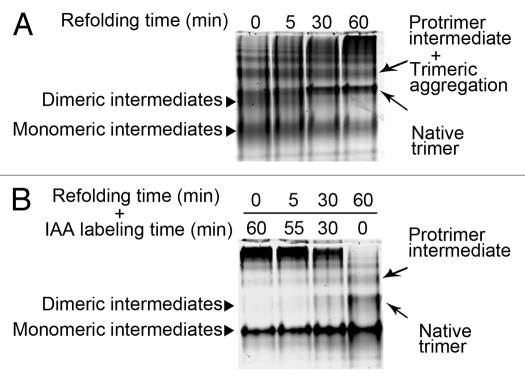
Elongated trimeric adhesins are a distinct class of proteins employed by phages and viruses to recognize and bind to their host cells, and by bacteria to bind to their target cells and tissues. The tailspikes of E. coli phage K1F and Bacillus phage Ø29 exhibit auto-chaperone activity in their trimeric C-terminal domains. The P22 tailspike is structurally homologous to those adhesins. Though there are no disulfide bonds or reactive cysteines in the native P22 tailspikes, a set of C-terminal cysteines are very reactive in partially folded intermediates, implying an unusual local conformation in the domain. This is likely to be involved in the auto-chaperone function. We examined the unusual reactivity of C-terminal tailspike cysteines during folding and assembly as a potential reporter of auto-chaperone function. Reaction with IAA blocked productive refolding in vitro, but not off-pathway aggregation. Two-dimensional PAGE revealed that the predominant intermediate exhibiting reactive cysteine side chains was a partially folded monomer. Treatment with reducing reagent promoted native trimer formation from these species, consistent with transient disulfide bonds in the auto-chaperone domain. Limited enzymatic digestion and mass spectrometry of folding and assembly intermediates indicated that the C-terminal domain was compact in the protrimer species. These results indicate that the C-terminal domain of the P22 tailspike folds itself and associates prior to formation of the protrimer intermediate, and not after, as previously proposed. The C-terminal cysteines and triple β-helix domains apparently provide the staging for the correct auto-chaperone domain formation, needed for alignment of P22 tailspike native trimer.
Figures
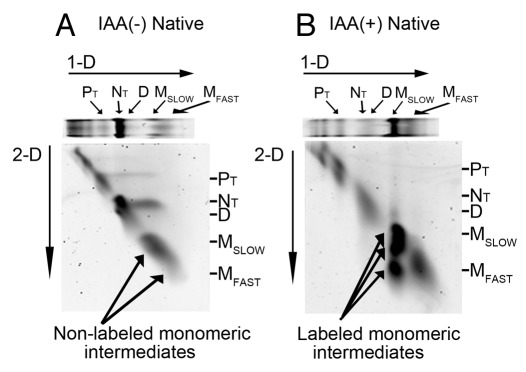
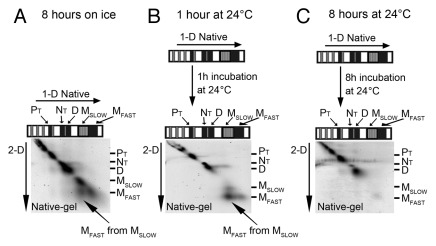

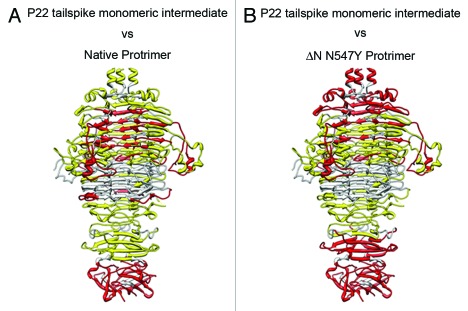

Similar articles
-
Characterization of the protrimer intermediate in the folding pathway of the interdigitated beta-helix tailspike protein.Biochemistry. 2002 Apr 23;41(16):5093-103. doi: 10.1021/bi0115582. Biochemistry. 2002. PMID: 11955057
-
Role for cysteine residues in the in vivo folding and assembly of the phage P22 tailspike.Protein Sci. 2001 Feb;10(2):397-410. doi: 10.1110/ps.34701. Protein Sci. 2001. PMID: 11266625 Free PMC article.
-
P22 tailspike trimer assembly is governed by interchain redox associations.Biochim Biophys Acta. 2004 Jul 1;1700(1):105-16. doi: 10.1016/j.bbapap.2004.04.001. Biochim Biophys Acta. 2004. PMID: 15210130
-
Stalled folding mutants in the triple beta-helix domain of the phage P22 tailspike adhesin.J Mol Biol. 2005 Dec 16;354(5):1103-17. doi: 10.1016/j.jmb.2005.10.007. Epub 2005 Oct 27. J Mol Biol. 2005. PMID: 16289113
-
There's a right way and a wrong way: in vivo and in vitro folding, misfolding and subunit assembly of the P22 tailspike.Structure. 1999 Jun 15;7(6):R131-9. doi: 10.1016/s0969-2126(99)80078-1. Structure. 1999. PMID: 10404587 Review.
Cited by
-
Structure of the Receptor-Binding Carboxy-Terminal Domain of the Bacteriophage T5 L-Shaped Tail Fibre with and without Its Intra-Molecular Chaperone.Viruses. 2015 Dec 8;7(12):6424-40. doi: 10.3390/v7122946. Viruses. 2015. PMID: 26670244 Free PMC article.
References
-
- Brennan MJ, David JL, Kenimer JG, Manclark CR. Lectin-like binding of pertussis toxin to a 165-kilodalton Chinese hamster ovary cell glycoprotein. J Biol Chem. 1988;263:4895–9. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources