

Transient B cell depletion or improved transgene expression by codon optimization promote tolerance to factor VIII in gene therapy
- PMID: 22655063
- PMCID: PMC3359994
- DOI: 10.1371/journal.pone.0037671
Transient B cell depletion or improved transgene expression by codon optimization promote tolerance to factor VIII in gene therapy
Abstract
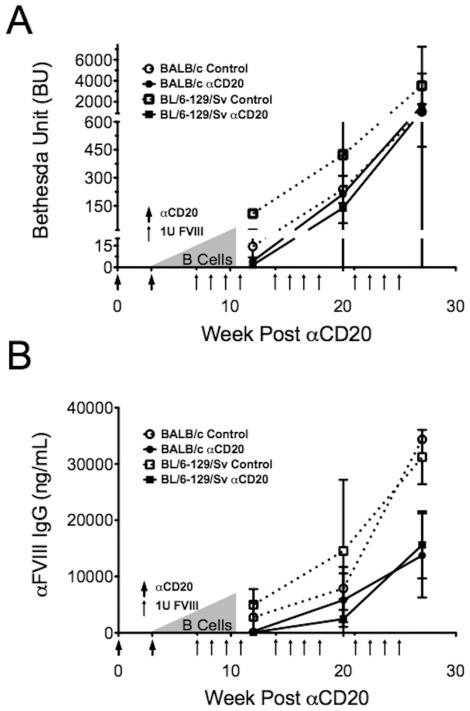
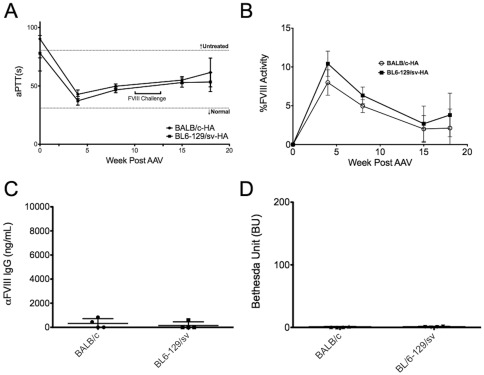
The major complication in the treatment of hemophilia A is the development of neutralizing antibodies (inhibitors) against factor VIII (FVIII). The current method for eradicating inhibitors, termed immune tolerance induction (ITI), is costly and protracted. Clinical protocols that prevent rather than treat inhibitors are not yet established. Liver-directed gene therapy hopes to achieve long-term correction of the disease while also inducing immune tolerance. We sought to investigate the use of adeno-associated viral (serotype 8) gene transfer to induce tolerance to human B domain deleted FVIII in hemophilia A mice. We administered an AAV8 vector with either human B domain deleted FVIII or a codon-optimized transgene, both under a liver-specific promoter to two strains of hemophilia A mice. Protein therapy or gene therapy was given either alone or in conjunction with anti-CD20 antibody-mediated B cell depletion. Gene therapy with a low-expressing vector resulted in sustained near-therapeutic expression. However, supplementary protein therapy revealed that gene transfer had sensitized mice to hFVIII in a high-responder strain but not in mice of a low-responding strain. This heightened response was ameliorated when gene therapy was delivered with anti-murine CD20 treatment. Transient B cell depletion prevented inhibitor formation in protein therapy, but failed to achieve a sustained hypo-responsiveness. Importantly, use of a codon-optimized hFVIII transgene resulted in sustained therapeutic expression and tolerance without a need for B cell depletion. Therefore, anti-CD20 may be beneficial in preventing vector-induced immune priming to FVIII, but higher levels of liver-restricted expression are preferred for tolerance.
Conflict of interest statement
Figures
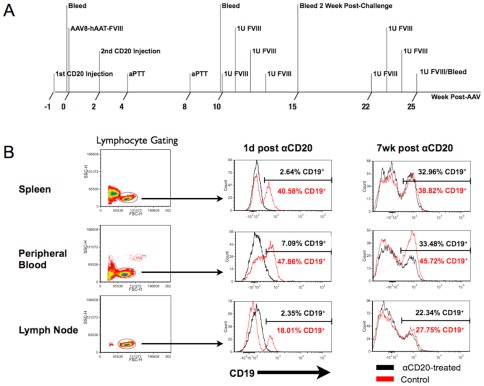
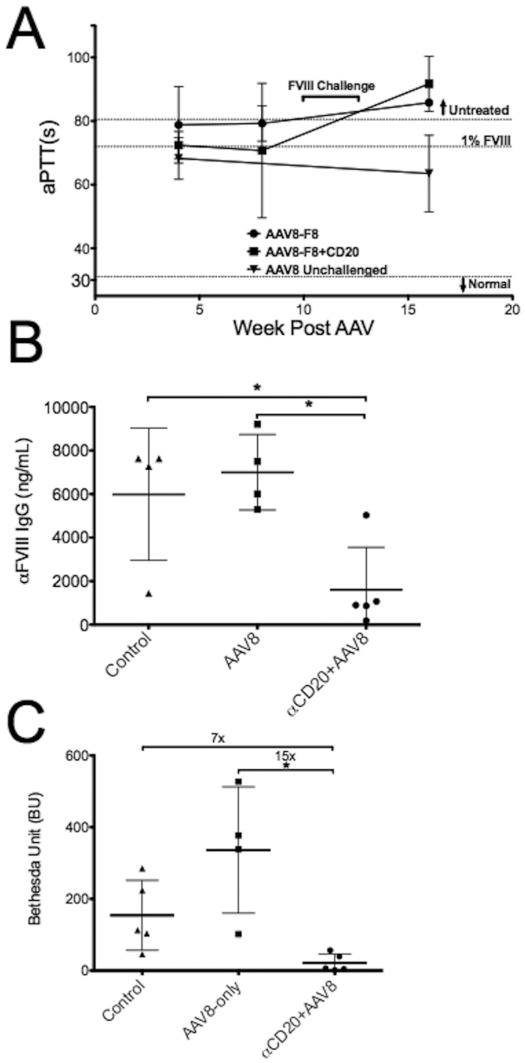
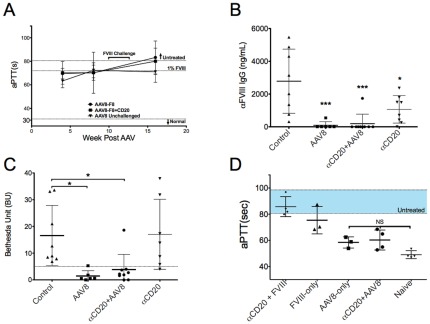
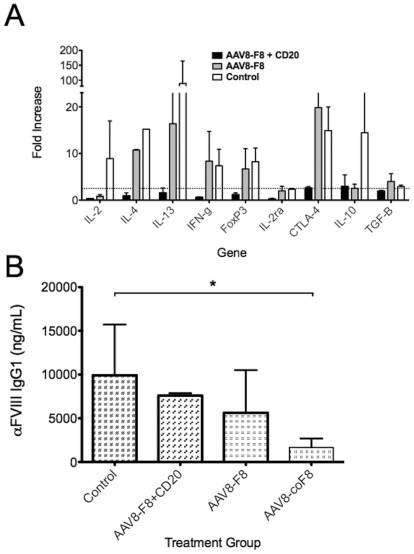
Similar articles
-
B Cell Depletion Eliminates FVIII Memory B Cells and Enhances AAV8-coF8 Immune Tolerance Induction When Combined With Rapamycin.Front Immunol. 2020 Jun 24;11:1293. doi: 10.3389/fimmu.2020.01293. eCollection 2020. Front Immunol. 2020. PMID: 32670285 Free PMC article.
-
Characterization of Adeno-Associated Viral Vector-Mediated Human Factor VIII Gene Therapy in Hemophilia A Mice.Hum Gene Ther. 2017 May;28(5):392-402. doi: 10.1089/hum.2016.128. Epub 2017 Jan 5. Hum Gene Ther. 2017. PMID: 28056565
-
Combination therapy for inhibitor reversal in haemophilia A using monoclonal anti-CD20 and rapamycin.Thromb Haemost. 2017 Jan 5;117(1):33-43. doi: 10.1160/TH16-05-0404. Epub 2016 Sep 29. Thromb Haemost. 2017. PMID: 27683758 Free PMC article.
-
Translational Potential of Immune Tolerance Induction by AAV Liver-Directed Factor VIII Gene Therapy for Hemophilia A.Front Immunol. 2020 Apr 28;11:618. doi: 10.3389/fimmu.2020.00618. eCollection 2020. Front Immunol. 2020. PMID: 32425925 Free PMC article. Review.
-
The Immune Response to the fVIII Gene Therapy in Preclinical Models.Front Immunol. 2020 Apr 15;11:494. doi: 10.3389/fimmu.2020.00494. eCollection 2020. Front Immunol. 2020. PMID: 32351497 Free PMC article. Review.
Cited by
-
IL-15 blockade and rapamycin rescue multifactorial loss of factor VIII from AAV-transduced hepatocytes in hemophilia A mice.Mol Ther. 2022 Dec 7;30(12):3552-3569. doi: 10.1016/j.ymthe.2022.07.005. Epub 2022 Jul 12. Mol Ther. 2022. PMID: 35821634 Free PMC article.
-
Complexity of immune responses to AAV transgene products - Example of factor IX.Cell Immunol. 2019 Aug;342:103658. doi: 10.1016/j.cellimm.2017.05.006. Epub 2017 May 29. Cell Immunol. 2019. PMID: 28645365 Free PMC article. Review.
-
piggyBac-mediated phenotypic correction of factor VIII deficiency.Mol Ther Methods Clin Dev. 2014 Sep 10;1:14042. doi: 10.1038/mtm.2014.42. eCollection 2014. Mol Ther Methods Clin Dev. 2014. PMID: 26015980 Free PMC article.
-
Targeting Antigen-Specific B Cells Using Antigen-Expressing Transduced Regulatory T Cells.J Immunol. 2018 Sep 1;201(5):1434-1441. doi: 10.4049/jimmunol.1701800. Epub 2018 Jul 18. J Immunol. 2018. PMID: 30021767 Free PMC article.
-
Emerging Issues in AAV-Mediated In Vivo Gene Therapy.Mol Ther Methods Clin Dev. 2017 Dec 1;8:87-104. doi: 10.1016/j.omtm.2017.11.007. eCollection 2018 Mar 16. Mol Ther Methods Clin Dev. 2017. PMID: 29326962 Free PMC article. Review.
References
-
- Di Michele DM. Immune tolerance induction in haemophilia: evidence and the way forward. J Thromb Haemost. 2011;9(Suppl 1):216–225. - PubMed
-
- Odeyemi IA, Dano AM. Optimising immune tolerance induction strategies in the management of haemophilia patients with inhibitors: a cost-minimisation analysis. Curr Med Res Opin. 2009;25:239–250. - PubMed
-
- Verweij CL, Vosslamber S. New insight in the mechanism of action of rituximab: the interferon signature towards personalized medicine. Discov Med. 2011;12:229–236. - PubMed
-
- Aleem A, Saidu A, Abdulkarim H, Al-Diab AR, Al-Sagheer A, et al. Rituximab as a single agent in the management of adult patients with haemophilia A and inhibitors: marked reduction in inhibitor level and clinical improvement in bleeding but failure to eradicate the inhibitor. Haemophilia. 2009;15:210–216. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01 DK58327/DK/NIDDK NIH HHS/United States
- R01 HL51390/HL/NHLBI NIH HHS/United States
- UL1 TR000064/TR/NCATS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- P01 DK058327/DK/NIDDK NIH HHS/United States
- P01 HL59412/HL/NHLBI NIH HHS/United States
- R01 HL073838/HL/NHLBI NIH HHS/United States
- P01 HL059412/HL/NHLBI NIH HHS/United States
- P01 HL078810/HL/NHLBI NIH HHS/United States
- R01 HL109442/HL/NHLBI NIH HHS/United States
- P01 HD078810/HD/NICHD NIH HHS/United States
- R01 AI051390/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
