

The reverse, but coordinated, roles of Tor2 (TORC1) and Tor1 (TORC2) kinases for growth, cell cycle and separase-mediated mitosis in Schizosaccharomyces pombe
- PMID: 22645648
- PMCID: PMC3352084
- DOI: 10.1098/rsob.110007
The reverse, but coordinated, roles of Tor2 (TORC1) and Tor1 (TORC2) kinases for growth, cell cycle and separase-mediated mitosis in Schizosaccharomyces pombe
Abstract
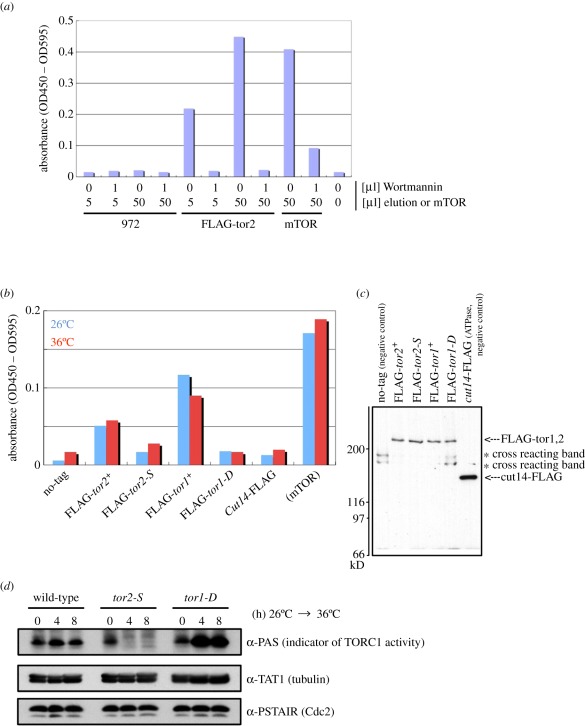
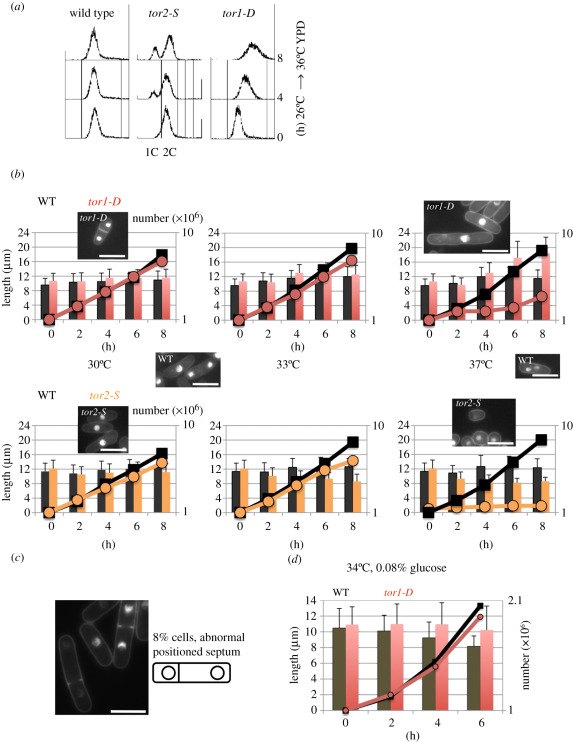
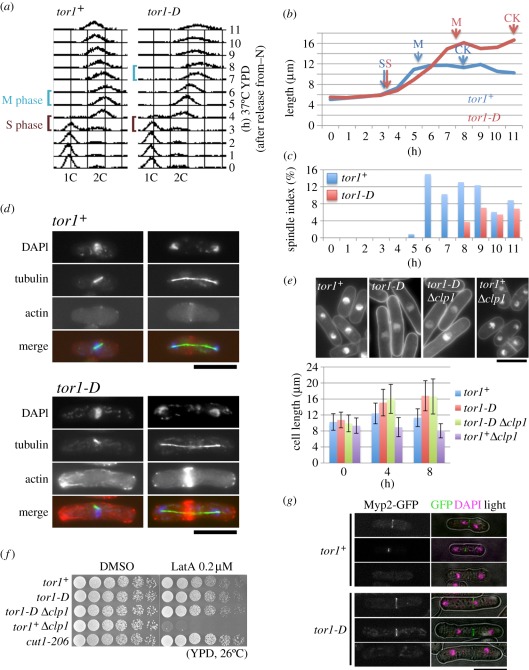
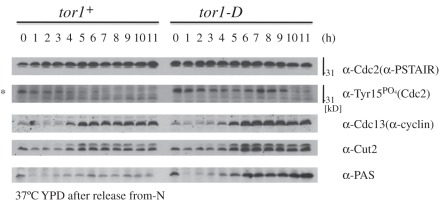
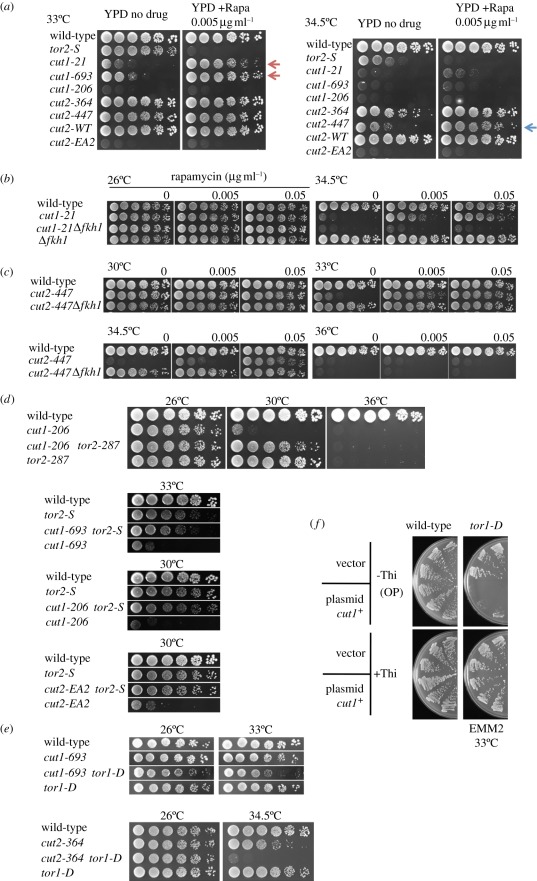
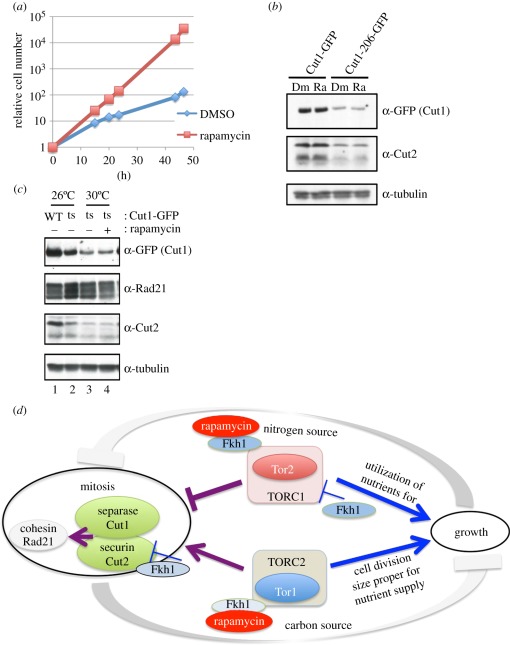
Target of rapamycin complexes (TORCs), which are vital for nutrient utilization, contain a catalytic subunit with the phosphatidyl inositol kinase-related kinase (PIKK) motif. TORC1 is required for cell growth, while the functions of TORC2 are less well understood. We show here that the fission yeast Schizosaccharomyces pombe TORC2 has a cell cycle role through determining the proper timing of Cdc2 Tyr15 dephosphorylation and the cell size under limited glucose, whereas TORC1 restrains mitosis and opposes securin-separase, which are essential for chromosome segregation. These results were obtained using the previously isolated TORC1 mutant tor2-L2048S in the phosphatidyl inositol kinase (PIK) domain and a new TORC2 mutant tor1-L2045D, which harbours a mutation in the same site. While mutated TORC1 and TORC2 displayed diminished kinase activity and FKBP12/Fkh1-dependent rapamycin sensitivity, their phenotypes were nearly opposite in mitosis. Premature mitosis and the G2-M delay occurred in TORC1 and TORC2 mutants, respectively. Surprisingly, separase/cut1-securin/cut2 mutants were rescued by TORC1/tor2-L2048S mutation or rapamycin addition or even Fkh1 deletion, whereas these mutants showed synthetic defect with TORC2/tor1-L2045D. TORC1 and TORC2 coordinate growth, mitosis and cell size control, such as Wee1 and Cdc25 do for the entry into mitosis.
Keywords: target of rapamycin, rapamycin, Fkh1, Cdc2, separase.
Figures

Similar articles
-
Nutrient limitations alter cell division control and chromosome segregation through growth-related kinases and phosphatases.Philos Trans R Soc Lond B Biol Sci. 2011 Dec 27;366(1584):3508-20. doi: 10.1098/rstb.2011.0124. Philos Trans R Soc Lond B Biol Sci. 2011. PMID: 22084378 Free PMC article. Review.
-
Fission yeast Tor1 functions as part of TORC1 to control mitotic entry through the stress MAPK pathway following nutrient stress.J Cell Sci. 2009 Jun 1;122(Pt 11):1737-46. doi: 10.1242/jcs.049387. Epub 2009 May 5. J Cell Sci. 2009. PMID: 19417002
-
TOR complex 2 controls gene silencing, telomere length maintenance, and survival under DNA-damaging conditions.Mol Cell Biol. 2009 Aug;29(16):4584-94. doi: 10.1128/MCB.01879-08. Epub 2009 Jun 22. Mol Cell Biol. 2009. PMID: 19546237 Free PMC article.
-
Separase-mediated cleavage of cohesin at interphase is required for DNA repair.Nature. 2004 Aug 26;430(7003):1044-8. doi: 10.1038/nature02803. Nature. 2004. PMID: 15329725
-
TOR signaling in fission yeast.Crit Rev Biochem Mol Biol. 2008 Jul-Aug;43(4):277-83. doi: 10.1080/10409230802254911. Crit Rev Biochem Mol Biol. 2008. PMID: 18756382 Review.
Cited by
-
The Loss of Lam2 and Npr2-Npr3 Diminishes the Vacuolar Localization of Gtr1-Gtr2 and Disinhibits TORC1 Activity in Fission Yeast.PLoS One. 2016 May 26;11(5):e0156239. doi: 10.1371/journal.pone.0156239. eCollection 2016. PLoS One. 2016. PMID: 27227887 Free PMC article.
-
Phosphorylation of the TOR ATP binding domain by AGC kinase constitutes a novel mode of TOR inhibition.J Cell Biol. 2013 Nov 25;203(4):595-604. doi: 10.1083/jcb.201305103. Epub 2013 Nov 18. J Cell Biol. 2013. PMID: 24247430 Free PMC article.
-
The GATA Transcription Factor Gaf1 Represses tRNAs, Inhibits Growth, and Extends Chronological Lifespan Downstream of Fission Yeast TORC1.Cell Rep. 2020 Mar 10;30(10):3240-3249.e4. doi: 10.1016/j.celrep.2020.02.058. Cell Rep. 2020. PMID: 32160533 Free PMC article.
-
Synergistic roles of the phospholipase B homolog Plb1 and the cAMP-dependent protein kinase Pka1 in the hypertonic stress response of Schizosaccharomyces pombe.Curr Genet. 2022 Dec;68(5-6):661-674. doi: 10.1007/s00294-022-01253-z. Epub 2022 Sep 16. Curr Genet. 2022. PMID: 36112198
-
Systematic screen for mutants resistant to TORC1 inhibition in fission yeast reveals genes involved in cellular ageing and growth.Biol Open. 2014 Feb 15;3(2):161-71. doi: 10.1242/bio.20147245. Biol Open. 2014. PMID: 24463365 Free PMC article.
References
-
- Schmelzle T, Hall MN. 2000. TOR, a central controller of cell growth. Cell 103, 253–26210.1016/S0092-8674(00)00117-3 (doi:10.1016/S0092-8674(00)00117-3) - DOI - DOI - PubMed
-
- Wullschleger S, Loewith R, Hall MN. 2006. TOR signaling in growth and metabolism. Cell 124, 471–48410.1016/j.cell.2006.01.016 (doi:10.1016/j.cell.2006.01.016) - DOI - DOI - PubMed
-
- Martel RR, Klicius J, Galet S. 1977. Inhibition of the immune response by rapamycin, a new antifungal antibiotic. Can. J. Physiol. Pharmacol. 55, 48–5110.1139/y77-007 (doi:10.1139/y77-007) - DOI - DOI - PubMed
-
- Sehgal SN, Baker H, Vezina C. 1975. Rapamycin (AY-22,989), a new antifungal antibiotic. II. Fermentation, isolation and characterization. J. Antibiotics 28, 727–732 - PubMed
-
- Zheng XF, Florentino D, Chen J, Crabtree GR, Schreiber SL. 1995. TOR kinase domains are required for two distinct functions, only one of which is inhibited by rapamycin. Cell 82, 121–13010.1016/0092-8674(95)90058-6 (doi:10.1016/0092-8674(95)90058-6) - DOI - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
