

Extracellular cyclophilin-A stimulates ERK1/2 phosphorylation in a cell-dependent manner but broadly stimulates nuclear factor kappa B
- PMID: 22631225
- PMCID: PMC3390265
- DOI: 10.1186/1475-2867-12-19
Extracellular cyclophilin-A stimulates ERK1/2 phosphorylation in a cell-dependent manner but broadly stimulates nuclear factor kappa B
Abstract
Background: Although the peptidyl-prolyl isomerase, cyclophilin-A (peptidyl-prolyl isomerase, PPIA), has been studied for decades in the context of its intracellular functions, its extracellular roles as a major contributor to both inflammation and multiple cancers have more recently emerged. A wide range of activities have been ascribed to extracellular PPIA that include induction of cytokine and matrix metalloproteinase (MMP) secretion, which potentially underlie its roles in inflammation and tumorigenesis. However, there have been conflicting reports as to which particular signaling events are under extracellular PPIA regulation, which may be due to either cell-dependent responses and/or the use of commercial preparations recently shown to be highly impure.
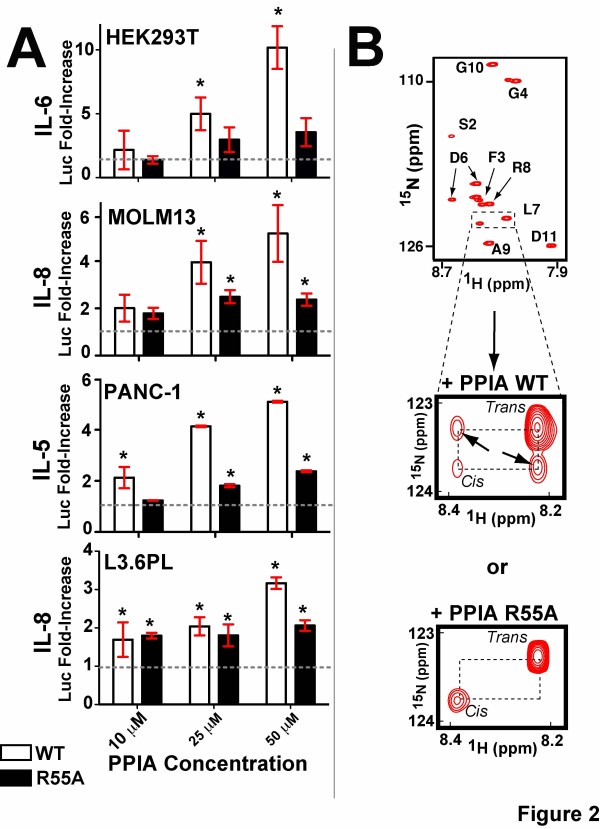
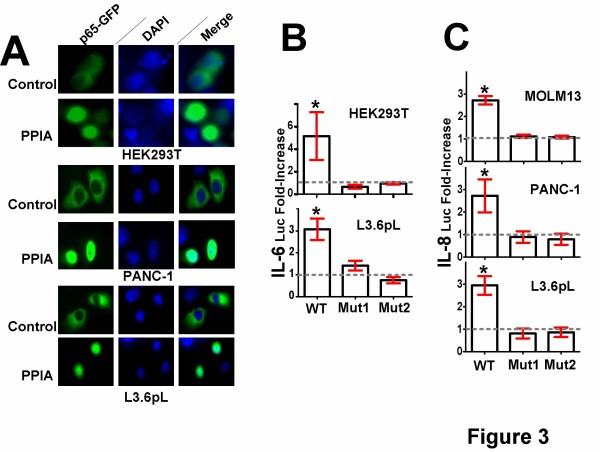
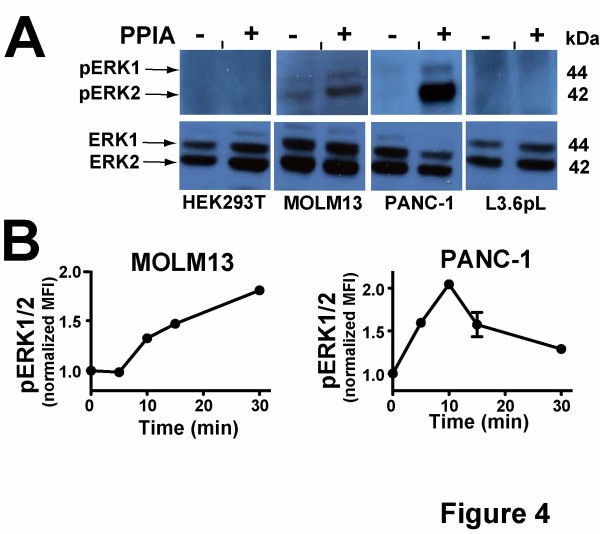
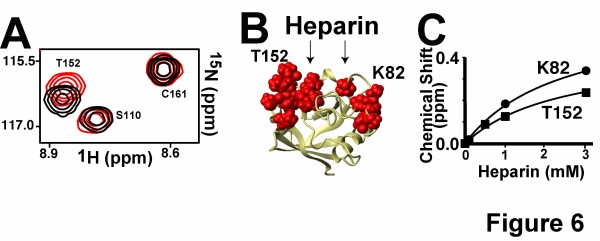
Methods: We have produced and validated the purity of recombinant PPIA in order to subject it to a comparative analysis between different cell types. Specifically, we have used a combination of multiple methods such as luciferase reporter screens, translocation assays, phosphorylation assays, and nuclear magnetic resonance to compare extracellular PPIA activities in several different cell lines that included epithelial and monocytic cells.
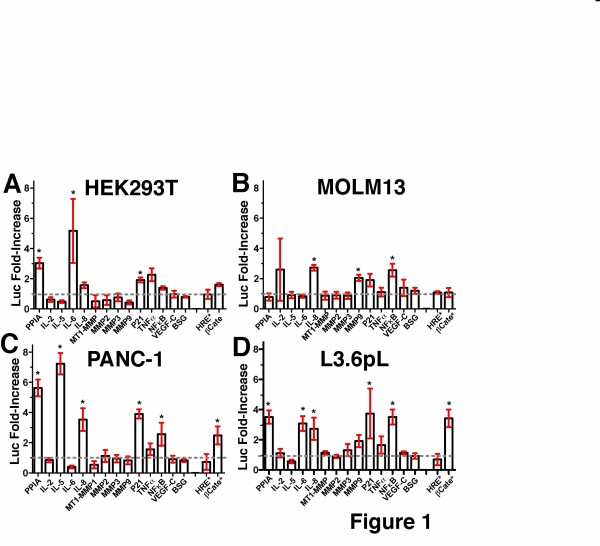
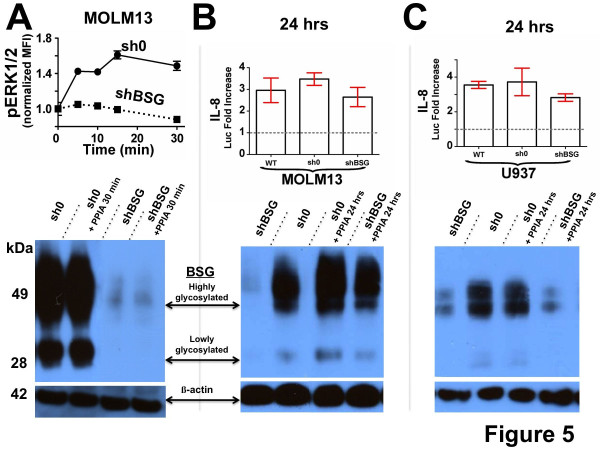
Results: Our findings have revealed that extracellular PPIA activity is cell type-dependent and that PPIA signals via multiple cellular receptors beyond the single transmembrane receptor previously identified, Extracellular Matrix MetalloPRoteinase Inducer (EMMPRIN). Finally, while our studies provide important insight into the cell-specific responses, they also indicate that there are consistent responses such as nuclear factor kappa B (NFκB) signaling induced in all cell lines tested.
Conclusions: We conclude that although extracellular PPIA activates several common pathways, it also targets different receptors in different cell types, resulting in a complex, integrated signaling network that is cell type-specific.
Figures
Similar articles
-
Targeting Extracellular Cyclophilin A Reduces Neuroinflammation and Extends Survival in a Mouse Model of Amyotrophic Lateral Sclerosis.J Neurosci. 2017 Feb 8;37(6):1413-1427. doi: 10.1523/JNEUROSCI.2462-16.2016. Epub 2016 Dec 23. J Neurosci. 2017. PMID: 28011744 Free PMC article.
-
Extracellular matrix metalloproteinase inducer (CD147) is a novel receptor on platelets, activates platelets, and augments nuclear factor kappaB-dependent inflammation in monocytes.Circ Res. 2008 Feb 15;102(3):302-9. doi: 10.1161/CIRCRESAHA.107.157990. Epub 2007 Nov 29. Circ Res. 2008. PMID: 18048771
-
PpiA, a surface PPIase of the cyclophilin family in Lactococcus lactis.PLoS One. 2012;7(3):e33516. doi: 10.1371/journal.pone.0033516. Epub 2012 Mar 19. PLoS One. 2012. PMID: 22442694 Free PMC article.
-
Osteopontin: it's role in regulation of cell motility and nuclear factor kappa B-mediated urokinase type plasminogen activator expression.IUBMB Life. 2005 Jun;57(6):441-7. doi: 10.1080/15216540500159424. IUBMB Life. 2005. PMID: 16012053 Review.
-
Regulation and Role of Store-Operated Ca2+ Entry in Cellular Proliferation.In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 12. In: Kozak JA, Putney JW Jr, editors. Calcium Entry Channels in Non-Excitable Cells. Boca Raton (FL): CRC Press/Taylor & Francis; 2018. Chapter 12. PMID: 30299656 Free Books & Documents. Review.
Cited by
-
1H, 13C, and 15N backbone and side chain resonance assignments of thermophilic Geobacillus kaustophilus cyclophilin-A.Biomol NMR Assign. 2014 Apr;8(1):23-7. doi: 10.1007/s12104-012-9445-3. Epub 2012 Nov 9. Biomol NMR Assign. 2014. PMID: 23138858 Free PMC article.
-
Potential Alternative Receptors for SARS-CoV-2-Induced Kidney Damage: TLR-4, KIM-1/TIM-1, and CD147.Front Biosci (Landmark Ed). 2024 Jan 12;29(1):8. doi: 10.31083/j.fbl2901008. Front Biosci (Landmark Ed). 2024. PMID: 38287815 Free PMC article. Review.
-
Extracellular and Intracellular Cyclophilin A, Native and Post-Translationally Modified, Show Diverse and Specific Pathological Roles in Diseases.Arterioscler Thromb Vasc Biol. 2018 May;38(5):986-993. doi: 10.1161/ATVBAHA.117.310661. Epub 2018 Mar 29. Arterioscler Thromb Vasc Biol. 2018. PMID: 29599134 Free PMC article. Review.
-
Extracellular cyclophilins in health and disease.Biochim Biophys Acta. 2015 Oct;1850(10):2087-95. doi: 10.1016/j.bbagen.2014.11.013. Epub 2014 Nov 18. Biochim Biophys Acta. 2015. PMID: 25445705 Free PMC article. Review.
-
The roles of CD147 and/or cyclophilin A in kidney diseases.Mediators Inflamm. 2014;2014:728673. doi: 10.1155/2014/728673. Epub 2014 Dec 17. Mediators Inflamm. 2014. PMID: 25580061 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
