

HPS4/SABRE regulates plant responses to phosphate starvation through antagonistic interaction with ethylene signalling
- PMID: 22615140
- PMCID: PMC3421987
- DOI: 10.1093/jxb/ers131
HPS4/SABRE regulates plant responses to phosphate starvation through antagonistic interaction with ethylene signalling
Abstract
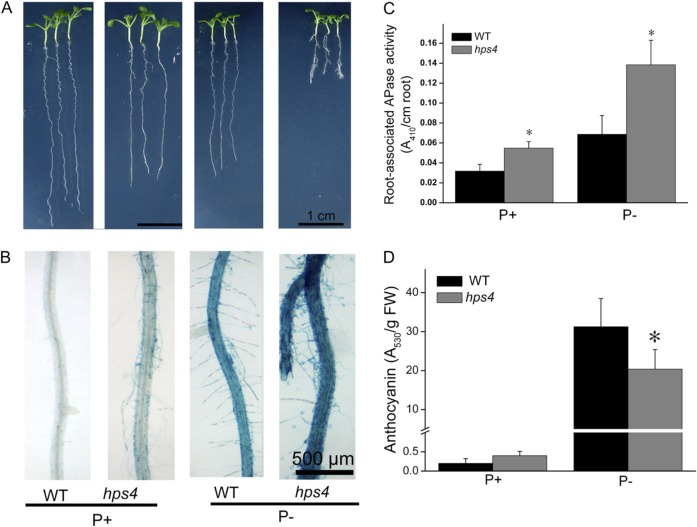
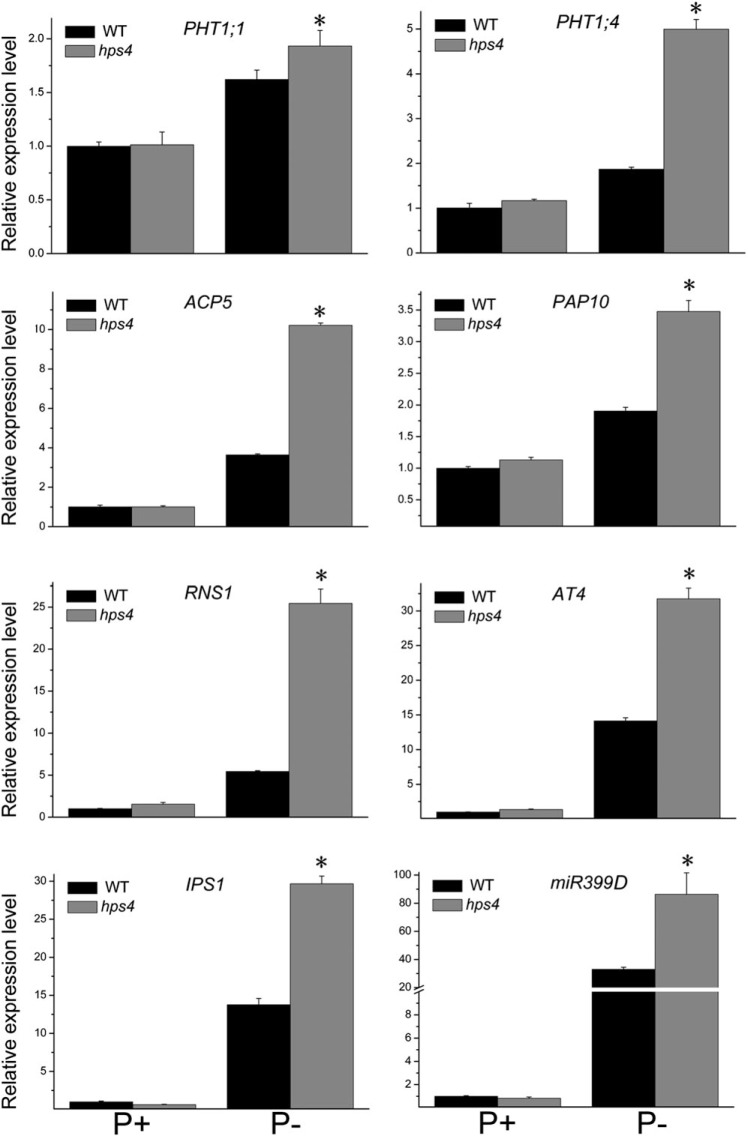
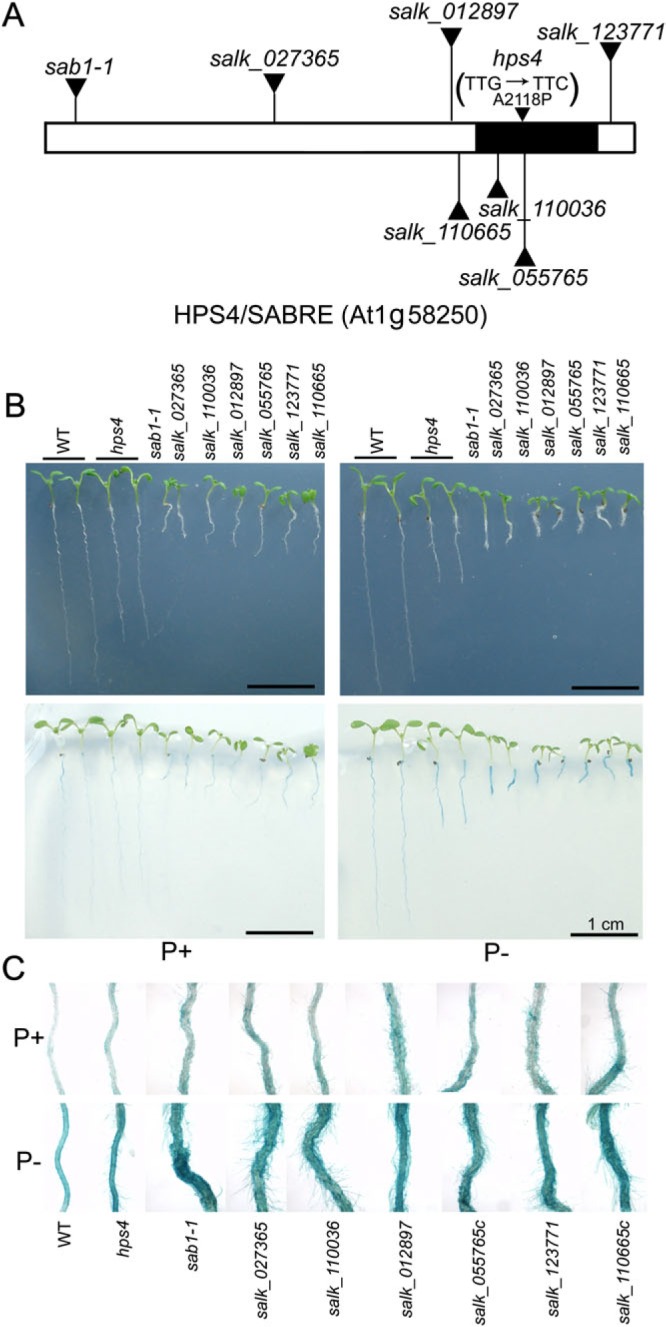
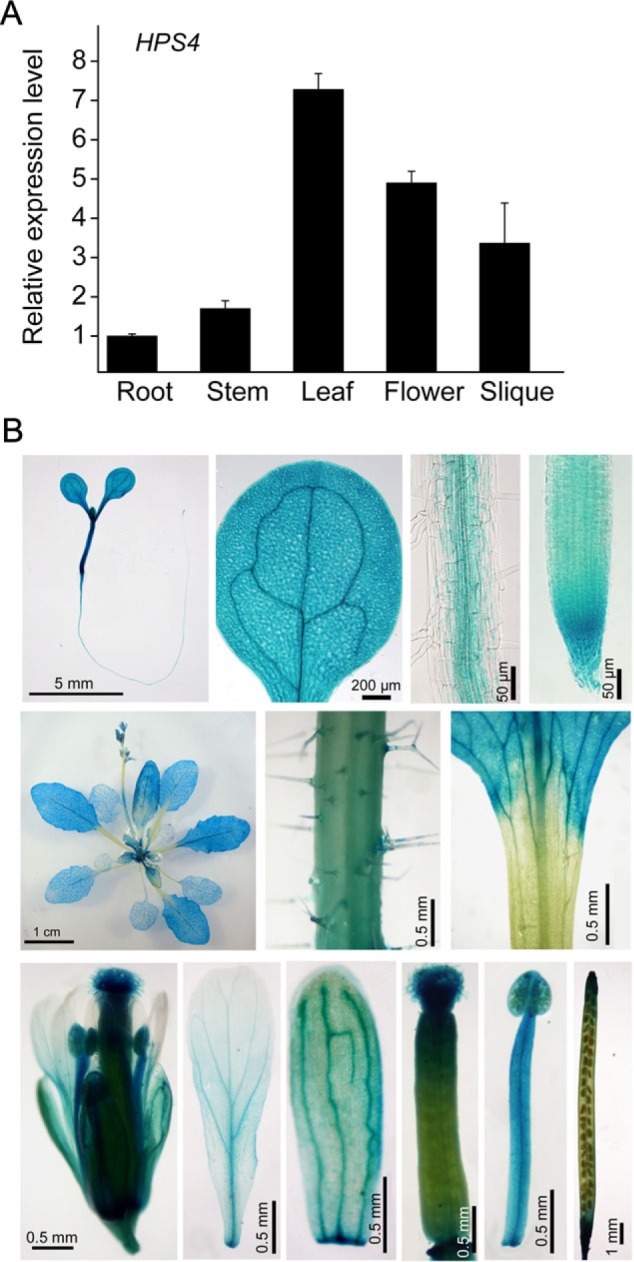
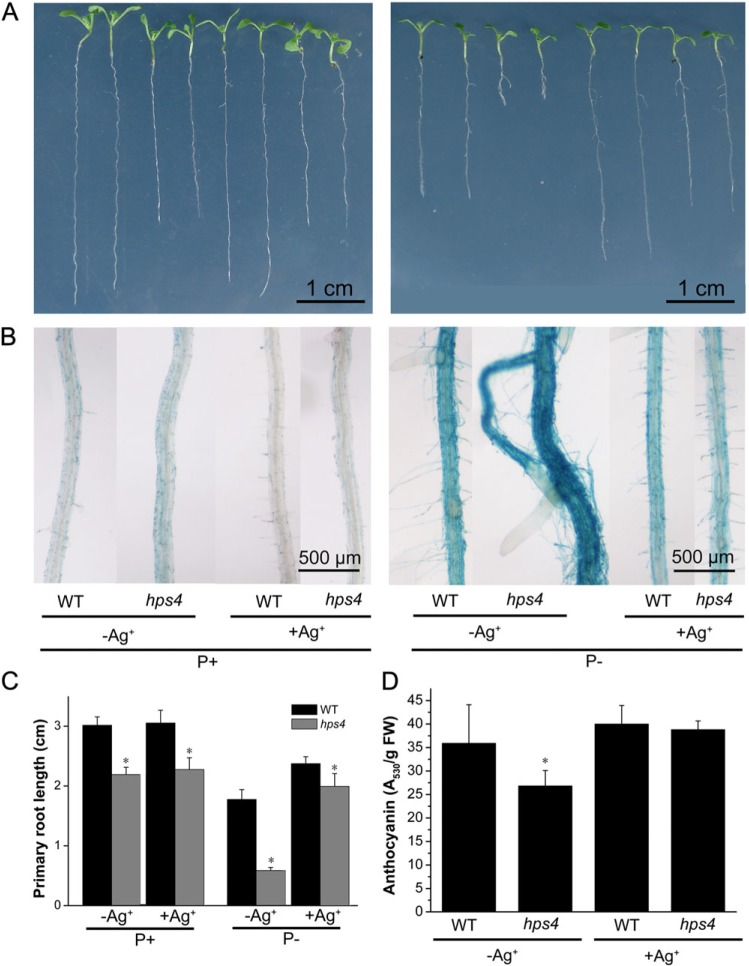
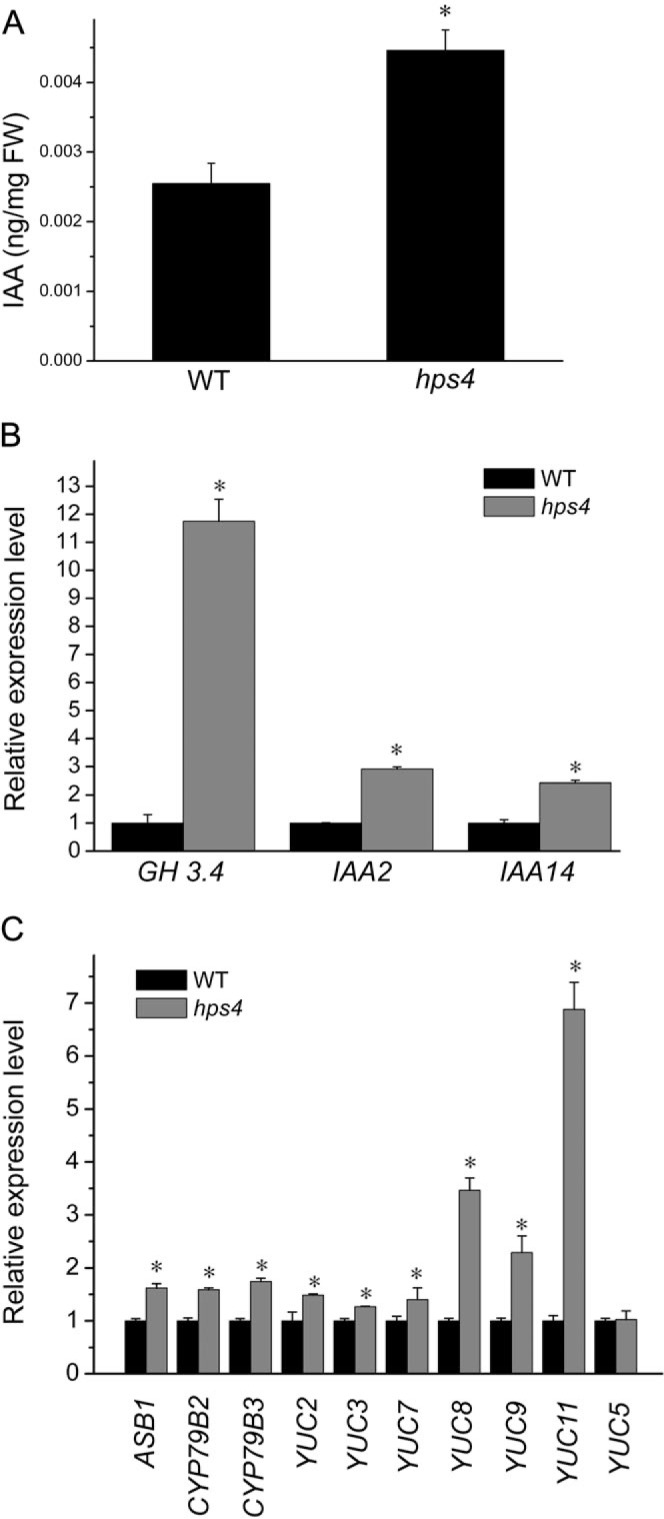
The phytohormone ethylene plays important roles in regulating plant responses to phosphate (Pi) starvation. To date, however, no molecular components have been identified that interact with ethylene signalling in regulating such responses. In this work, an Arabidopsis mutant, hps4, was characterized that exhibits enhanced responses to Pi starvation, including increased inhibition of primary root growth, enhanced expression of Pi starvation-induced genes, and overproduction of root-associated acid phosphatases. Molecular cloning indicated that hps4 is a new allele of SABRE, which was previously identified as an important regulator of cell expansion in Arabidopsis. HPS4/SABRE antagonistically interacts with ethylene signalling to regulate plant responses to Pi starvation. Furthermore, it is shown that Pi-starved hps4 mutants accumulate more auxin in their root tips than the wild type, which may explain the increased inhibition of their primary root growth when grown under Pi deficiency.
Figures
Similar articles
-
The Arabidopsis gene hypersensitive to phosphate starvation 3 encodes ethylene overproduction 1.Plant Cell Physiol. 2012 Jun;53(6):1093-105. doi: 10.1093/pcp/pcs072. Epub 2012 May 22. Plant Cell Physiol. 2012. PMID: 22623414
-
Ethylene signalling is involved in regulation of phosphate starvation-induced gene expression and production of acid phosphatases and anthocyanin in Arabidopsis.New Phytol. 2011 Mar;189(4):1084-1095. doi: 10.1111/j.1469-8137.2010.03555.x. Epub 2010 Nov 30. New Phytol. 2011. PMID: 21118263
-
A major root-associated acid phosphatase in Arabidopsis, AtPAP10, is regulated by both local and systemic signals under phosphate starvation.J Exp Bot. 2014 Dec;65(22):6577-88. doi: 10.1093/jxb/eru377. Epub 2014 Sep 22. J Exp Bot. 2014. PMID: 25246445 Free PMC article.
-
Adaptation to Phosphate Scarcity: Tips from Arabidopsis Roots.Trends Plant Sci. 2018 Aug;23(8):721-730. doi: 10.1016/j.tplants.2018.04.006. Epub 2018 May 12. Trends Plant Sci. 2018. PMID: 29764728 Review.
-
Ethylene's role in phosphate starvation signaling: more than just a root growth regulator.Plant Cell Physiol. 2012 Feb;53(2):277-86. doi: 10.1093/pcp/pcr186. Epub 2011 Dec 22. Plant Cell Physiol. 2012. PMID: 22199374 Review.
Cited by
-
The THO/TREX Complex Active in miRNA Biogenesis Negatively Regulates Root-Associated Acid Phosphatase Activity Induced by Phosphate Starvation.Plant Physiol. 2016 Aug;171(4):2841-53. doi: 10.1104/pp.16.00680. Epub 2016 Jun 21. Plant Physiol. 2016. PMID: 27329222 Free PMC article.
-
Interplay between Hormones and Several Abiotic Stress Conditions on Arabidopsis thaliana Primary Root Development.Cells. 2020 Dec 1;9(12):2576. doi: 10.3390/cells9122576. Cells. 2020. PMID: 33271980 Free PMC article. Review.
-
The Phosphorus-Iron Nexus: Decoding the Nutrients Interaction in Soil and Plant.Int J Mol Sci. 2024 Jun 26;25(13):6992. doi: 10.3390/ijms25136992. Int J Mol Sci. 2024. PMID: 39000100 Free PMC article. Review.
-
Comparative expression profiling reveals a role of the root apoplast in local phosphate response.BMC Plant Biol. 2016 Apr 28;16:106. doi: 10.1186/s12870-016-0790-8. BMC Plant Biol. 2016. PMID: 27121119 Free PMC article.
-
Adaptive strategies of plants to conserve internal phosphorus under P deficient condition to improve P utilization efficiency.Physiol Mol Biol Plants. 2022 Dec;28(11-12):1981-1993. doi: 10.1007/s12298-022-01255-8. Epub 2022 Dec 5. Physiol Mol Biol Plants. 2022. PMID: 36573147 Free PMC article. Review.
References
-
- Aeschbacher RA, Hauser M, Feldmann KA, Benfey P. The SABRE gene is required for normal cell expansion in Arabidopsis. Genes and Development. 1995;9:330–340. - PubMed
-
- Ames BN. Assay of inorganic phosphate, total phosphate and phosphatases. Methods in Enzymology. 1966;8:115–118.
-
- Bariola PA, Howard CJ, Taylor CB, Verburg MT, Jaglan VD, Green PJ. The Arabidopsis ribonuclease gene RNS1 is tightly controlled in response to phosphate limitation. The Plant Journal. 1994;6:673–685. - PubMed
-
- Borch K, Bouma TJ, Lynch JP, Brown KM. Ethylene: a regulator of root architectural responses to soil phosphorus availability. Plant, Cell and Environment. 1999;22:425–431.
-
- Boutin JP, Provot M, Roux L. Effect of cycloheximide and renewal of phosphorus supply on surface acid-phosphatase-activity of phosphorus deficient tomato roots. Physiologia Plantarum. 1981;51:353–360.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
