

Dualistic function of Daxx at centromeric and pericentromeric heterochromatin in normal and stress conditions
- PMID: 22572957
- PMCID: PMC3414404
- DOI: 10.4161/nucl.20180
Dualistic function of Daxx at centromeric and pericentromeric heterochromatin in normal and stress conditions
Abstract
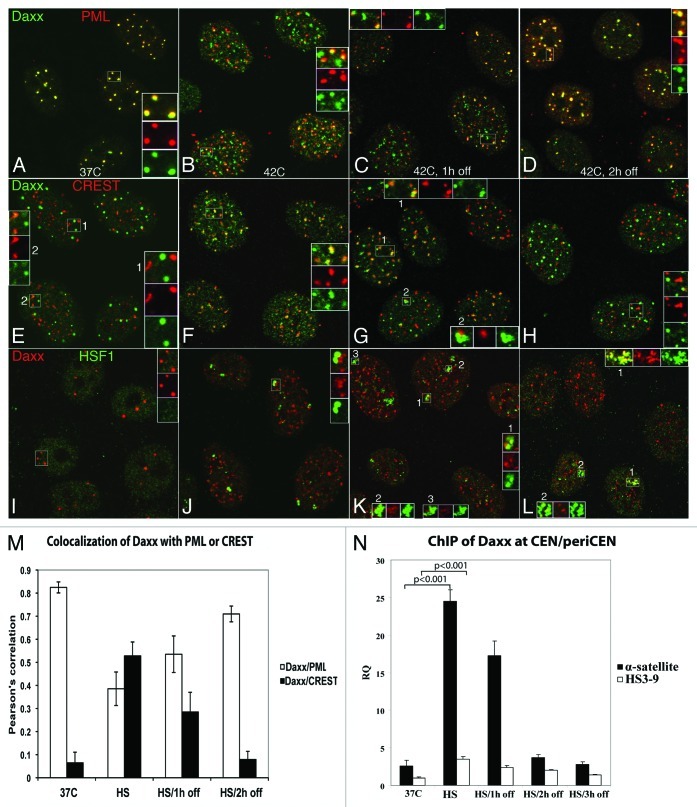
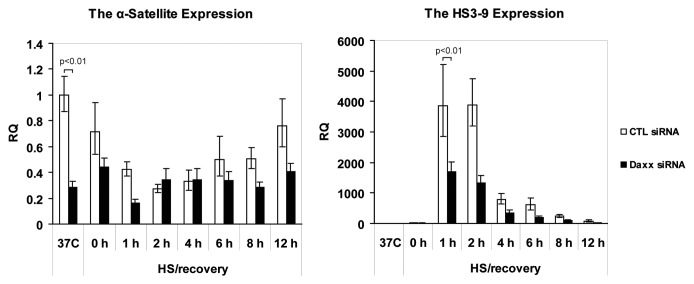
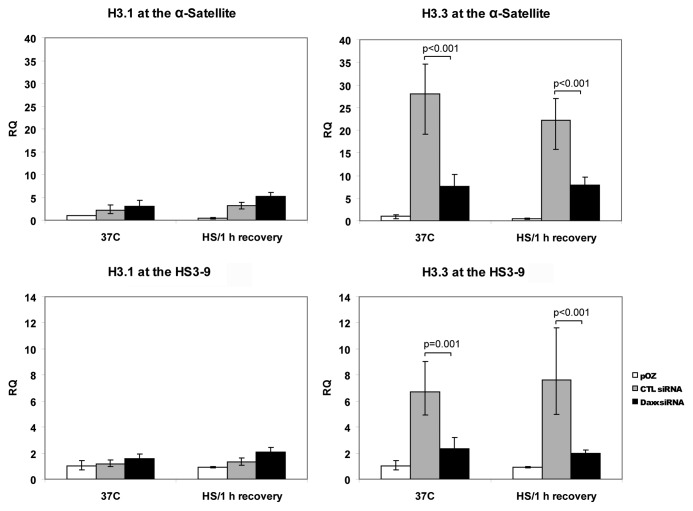
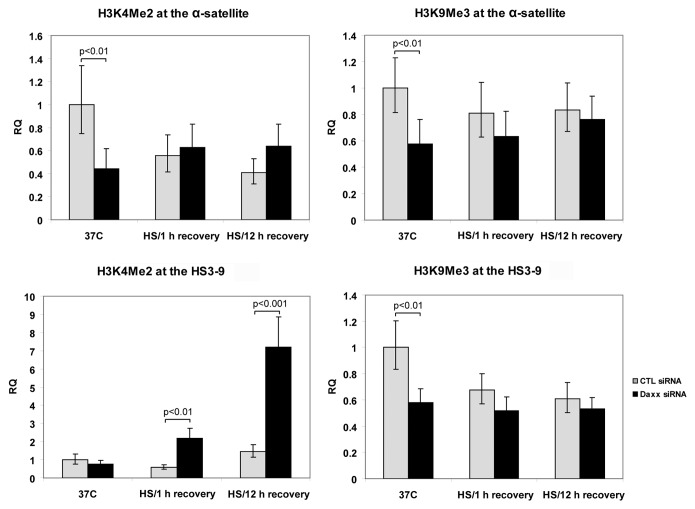
Nuclear structures ND10/PML NBs are linked to multiple processes, including the maintenance of intranuclear homeostasis by sequestering proteins into "nuclear depot." This function presumes release of proteins from PML NBs and their redistribution to the alternative, supposedly "active" locations, in response to the external stress application. To further investigate this nuclear depot function, we focused on the intranuclear distribution of protein Daxx that in normal conditions is mainly accumulated at PML NBs, and has a minor association with centromeres and pericentromeres (CEN/periCEN). Here we report that application of physiological Heat Shock (HS) changes this balance forcing very robust and reversible accumulation of Daxx on CEN/periCEN heterochromatin. Heterochromatin architecture is essential for the proper orchestration of nuclear processes, while transcription from this part of genome is required for its maintenance. To understand functional consequences of Daxx deposition at CEN/periCEN, we tested for Daxx-dependency of heterochromatin transcription. Depletion of Daxx reduces accumulation of CEN RNA in normal conditions and periCEN RNA after HS application. Searching for the mechanism of Daxx-dependent regulation of heterochromatin transcription, we found that depletion of Daxx decreases incorporation of transcription-associated histone H3 variant, H3.3, into both CEN and periCEN. Surprisingly, HS-induced deposition of Daxx does not further elevate incorporation of H3.3 into CEN/periCEN that remained steady during stress and recovery. Instead, depletion of Daxx leads to HS-induced changes in the balance of epigenetic modifications at heterochromatin, most dramatically elevating levels of active H3K4Me2 modification at periCEN. We propose dualistic function of Daxx-containing complexes at CEN/periCEN: (1) regulation of H3.3 loading in normal conditions and (2) protection of epigenetic status upon stress-induced accumulation, thus collectively guarding epigenetic identity of CEN/periCEN heterochromatin.
Figures
Similar articles
-
Dynamics of histone H3.3 deposition in proliferating and senescent cells reveals a DAXX-dependent targeting to PML-NBs important for pericentromeric heterochromatin organization.Cell Cycle. 2014;13(2):249-67. doi: 10.4161/cc.26988. Epub 2013 Nov 5. Cell Cycle. 2014. PMID: 24200965 Free PMC article.
-
CENP-B protects centromere chromatin integrity by facilitating histone deposition via the H3.3-specific chaperone Daxx.Epigenetics Chromatin. 2017 Dec 22;10(1):63. doi: 10.1186/s13072-017-0164-y. Epigenetics Chromatin. 2017. PMID: 29273057 Free PMC article.
-
The death-associated protein DAXX is a novel histone chaperone involved in the replication-independent deposition of H3.3.Genes Dev. 2010 Jun 15;24(12):1253-65. doi: 10.1101/gad.566910. Epub 2010 May 26. Genes Dev. 2010. PMID: 20504901 Free PMC article.
-
PML NBs (ND10) and Daxx: from nuclear structure to protein function.Front Biosci. 2008 May 1;13:7132-42. doi: 10.2741/3216. Front Biosci. 2008. PMID: 18508722 Review.
-
New players in heterochromatin silencing: histone variant H3.3 and the ATRX/DAXX chaperone.Nucleic Acids Res. 2016 Feb 29;44(4):1496-501. doi: 10.1093/nar/gkw012. Epub 2016 Jan 14. Nucleic Acids Res. 2016. PMID: 26773061 Free PMC article. Review.
Cited by
-
Nucleoli in embryos: a central structural platform for embryonic chromatin remodeling?Chromosome Res. 2019 Mar;27(1-2):129-140. doi: 10.1007/s10577-018-9590-3. Epub 2018 Nov 8. Chromosome Res. 2019. PMID: 30406864 Review.
-
Promyelocytic leukemia nuclear body-like structures can assemble in mouse oocytes.Biol Open. 2022 Jun 15;11(6):bio059130. doi: 10.1242/bio.059130. Epub 2022 Jun 6. Biol Open. 2022. PMID: 35579421 Free PMC article.
-
Centromeric cohesion failure invokes a conserved choreography of chromosomal mis-segregations in pancreatic neuroendocrine tumor.Genome Med. 2020 Apr 28;12(1):38. doi: 10.1186/s13073-020-00730-9. Genome Med. 2020. PMID: 32345369 Free PMC article.
-
Dynamics of histone H3.3 deposition in proliferating and senescent cells reveals a DAXX-dependent targeting to PML-NBs important for pericentromeric heterochromatin organization.Cell Cycle. 2014;13(2):249-67. doi: 10.4161/cc.26988. Epub 2013 Nov 5. Cell Cycle. 2014. PMID: 24200965 Free PMC article.
-
The histone chaperone DAXX maintains the structural organization of heterochromatin domains.Epigenetics Chromatin. 2015 Oct 21;8:44. doi: 10.1186/s13072-015-0036-2. eCollection 2015. Epigenetics Chromatin. 2015. PMID: 26500702 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous