

Links between metabolism and cancer
- PMID: 22549953
- PMCID: PMC3347786
- DOI: 10.1101/gad.189365.112
Links between metabolism and cancer
Abstract
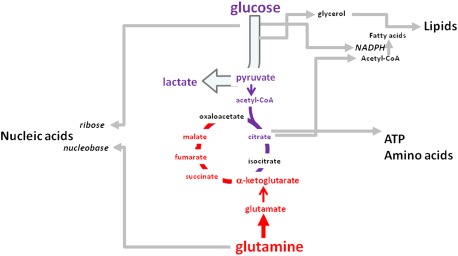
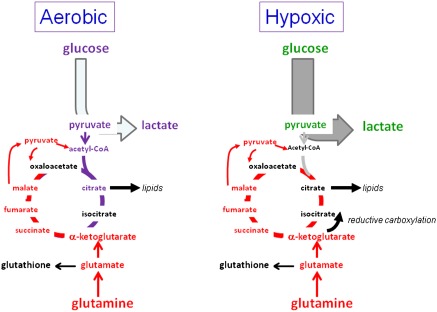
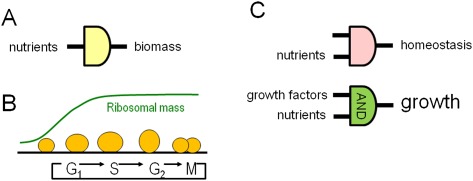
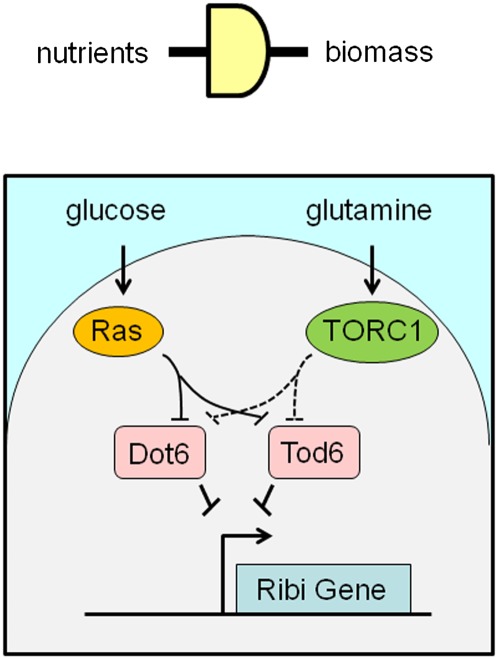
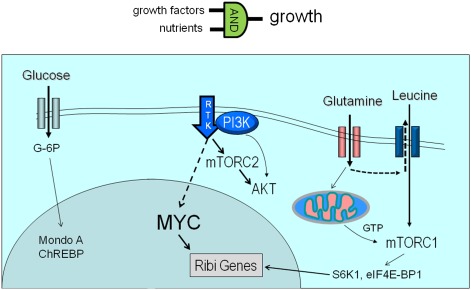
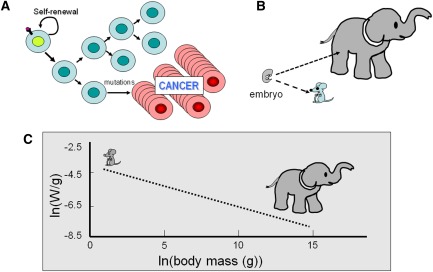
Metabolism generates oxygen radicals, which contribute to oncogenic mutations. Activated oncogenes and loss of tumor suppressors in turn alter metabolism and induce aerobic glycolysis. Aerobic glycolysis or the Warburg effect links the high rate of glucose fermentation to cancer. Together with glutamine, glucose via glycolysis provides the carbon skeletons, NADPH, and ATP to build new cancer cells, which persist in hypoxia that in turn rewires metabolic pathways for cell growth and survival. Excessive caloric intake is associated with an increased risk for cancers, while caloric restriction is protective, perhaps through clearance of mitochondria or mitophagy, thereby reducing oxidative stress. Hence, the links between metabolism and cancer are multifaceted, spanning from the low incidence of cancer in large mammals with low specific metabolic rates to altered cancer cell metabolism resulting from mutated enzymes or cancer genes.
Figures
Similar articles
-
The Warburg effect: essential part of metabolic reprogramming and central contributor to cancer progression.Int J Radiat Biol. 2019 Jul;95(7):912-919. doi: 10.1080/09553002.2019.1589653. Epub 2019 Mar 22. Int J Radiat Biol. 2019. PMID: 30822194 Review.
-
The control of the metabolic switch in cancers by oncogenes and tumor suppressor genes.Science. 2010 Dec 3;330(6009):1340-4. doi: 10.1126/science.1193494. Science. 2010. PMID: 21127244 Review.
-
Tumor microenvironment and metabolic synergy in breast cancers: critical importance of mitochondrial fuels and function.Semin Oncol. 2014 Apr;41(2):195-216. doi: 10.1053/j.seminoncol.2014.03.002. Epub 2014 Mar 5. Semin Oncol. 2014. PMID: 24787293 Review.
-
MYC-induced cancer cell energy metabolism and therapeutic opportunities.Clin Cancer Res. 2009 Nov 1;15(21):6479-83. doi: 10.1158/1078-0432.CCR-09-0889. Epub 2009 Oct 27. Clin Cancer Res. 2009. PMID: 19861459 Free PMC article. Review.
-
Mechanisms of Metabolic Reprogramming in Cancer Cells Supporting Enhanced Growth and Proliferation.Cells. 2021 Apr 29;10(5):1056. doi: 10.3390/cells10051056. Cells. 2021. PMID: 33946927 Free PMC article. Review.
Cited by
-
TMEFF2 and SARDH cooperate to modulate one-carbon metabolism and invasion of prostate cancer cells.Prostate. 2013 Oct;73(14):1561-75. doi: 10.1002/pros.22706. Epub 2013 Jul 3. Prostate. 2013. PMID: 23824605 Free PMC article.
-
NRF2-directed PRPS1 upregulation to promote the progression and metastasis of melanoma.Front Immunol. 2022 Sep 20;13:989263. doi: 10.3389/fimmu.2022.989263. eCollection 2022. Front Immunol. 2022. PMID: 36203561 Free PMC article.
-
Association of Irisin/FNDC5 with ERRα and PGC-1α Expression in NSCLC.Int J Mol Sci. 2022 Nov 17;23(22):14204. doi: 10.3390/ijms232214204. Int J Mol Sci. 2022. PMID: 36430689 Free PMC article.
-
Extracellular pH Modulates Neuroendocrine Prostate Cancer Cell Metabolism and Susceptibility to the Mitochondrial Inhibitor Niclosamide.PLoS One. 2016 Jul 20;11(7):e0159675. doi: 10.1371/journal.pone.0159675. eCollection 2016. PLoS One. 2016. PMID: 27438712 Free PMC article.
-
A link between cold environment and cancer.Tumour Biol. 2015 Aug;36(8):5953-64. doi: 10.1007/s13277-015-3270-0. Epub 2015 Mar 4. Tumour Biol. 2015. PMID: 25736923
References
-
- Bello-Fernandez C, Cleveland JL 1992. c-myc transactivates the ornithine decarboxylase gene. Curr Top Microbiol Immunol 182: 445–452 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources