

Human invariant natural killer T cells acquire transient innate responsiveness via histone H4 acetylation induced by weak TCR stimulation
- PMID: 22508835
- PMCID: PMC3348100
- DOI: 10.1084/jem.20111024
Human invariant natural killer T cells acquire transient innate responsiveness via histone H4 acetylation induced by weak TCR stimulation
Abstract
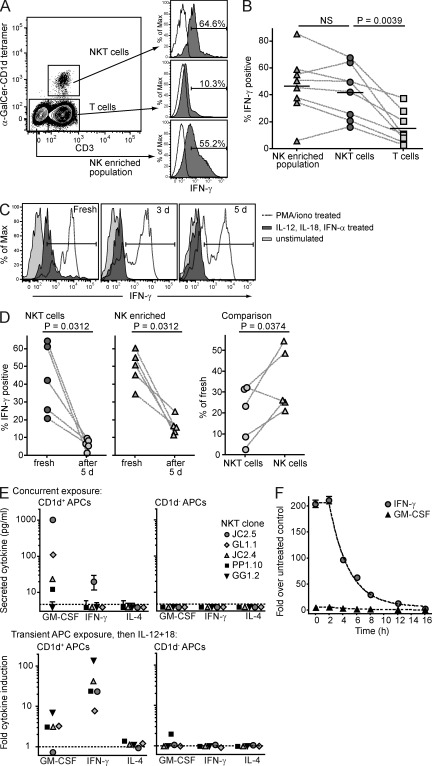
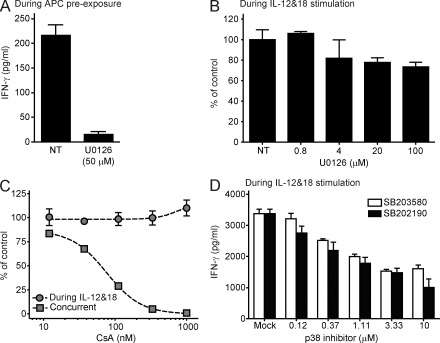
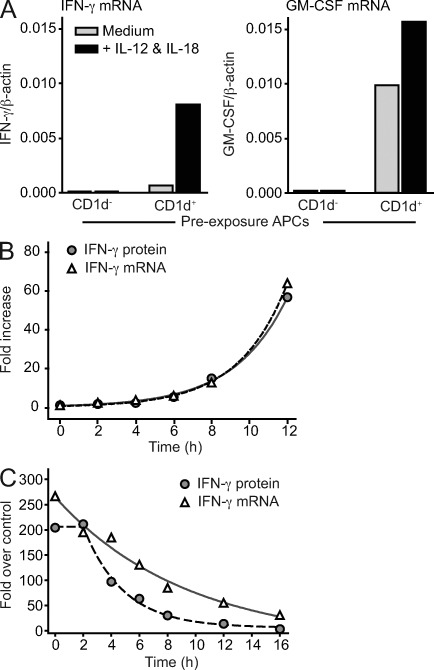
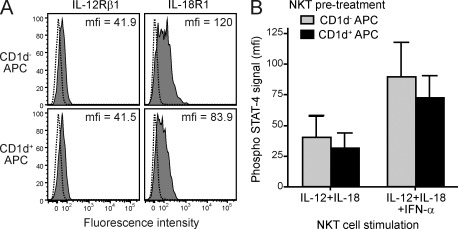
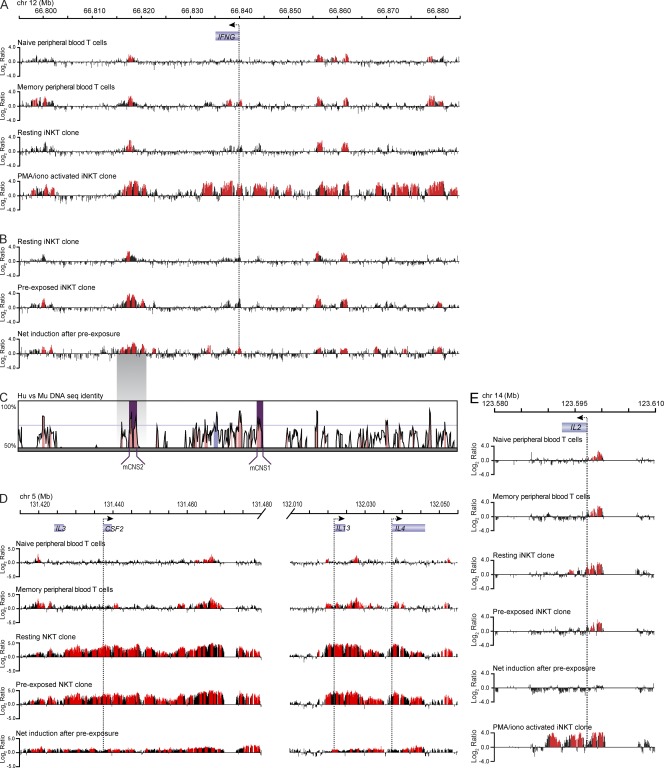
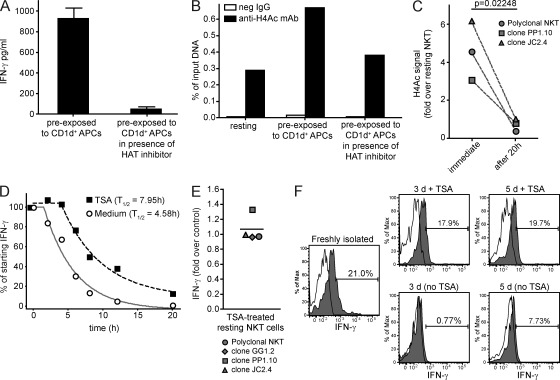
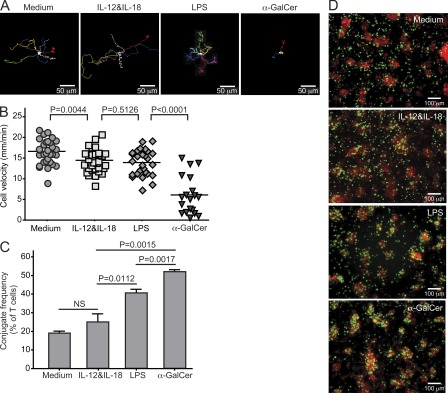
Invariant NKT cells (iNKT cells) are innate T lymphocytes that are thought to play an important role in producing an early burst of IFN-γ that promotes successful tumor immunosurveillance and antimicrobial immunity. The cellular activation processes underlying innate IFN-γ production remain poorly understood. We show here that weak T cell receptor (TCR) stimulation that does not directly activate iNKT cell IFN-γ messenger RNA transcription nevertheless induces histone H4 acetylation at specific regions near the IFNG gene locus. This renders the iNKT cells able to produce IFN-γ in an innate manner (i.e., not requiring concurrent TCR stimulation) upon exposure to IL-12 and IL-18. The iNKT cells retain the capacity for innate activation for hours to days after the initial weak TCR stimulation, although their innate responsiveness gradually declines as a function of histone deacetylation. These results explain how iNKT cells are able to mediate rapid innate IFN-γ secretion in a manner that does not require them to undergo permanent T(H1) differentiation. Moreover, our results also indicate that iNKT cell motility is maintained during activation by IL-12 and IL-18. Therefore, iNKT cells activated through this pathway can continue to migrate and may thus disseminate the IFN-γ that they produce, which may amplify its impact.
Figures
Similar articles
-
CD8 T cells in innate immune responses: using STAT4-dependent but antigen-independent pathways to gamma interferon during viral infection.mBio. 2014 Oct 21;5(5):e01978-14. doi: 10.1128/mBio.01978-14. mBio. 2014. PMID: 25336459 Free PMC article.
-
Transcriptional regulator Bhlhe40 works as a cofactor of T-bet in the regulation of IFN-γ production in iNKT cells.Proc Natl Acad Sci U S A. 2016 Jun 14;113(24):E3394-402. doi: 10.1073/pnas.1604178113. Epub 2016 May 25. Proc Natl Acad Sci U S A. 2016. PMID: 27226296 Free PMC article.
-
Combination of NKT14m and Low Dose IL-12 Promotes Invariant Natural Killer T Cell IFN-γ Production and Tumor Control.Int J Mol Sci. 2020 Jul 18;21(14):5085. doi: 10.3390/ijms21145085. Int J Mol Sci. 2020. PMID: 32708464 Free PMC article.
-
The Role of Invariant NKT in Autoimmune Liver Disease: Can Vitamin D Act as an Immunomodulator?Can J Gastroenterol Hepatol. 2018 Jun 26;2018:8197937. doi: 10.1155/2018/8197937. eCollection 2018. Can J Gastroenterol Hepatol. 2018. PMID: 30046564 Free PMC article. Review.
-
Functions of CD1d-Restricted Invariant Natural Killer T Cells in Antimicrobial Immunity and Potential Applications for Infection Control.Front Immunol. 2018 Jun 6;9:1266. doi: 10.3389/fimmu.2018.01266. eCollection 2018. Front Immunol. 2018. PMID: 29928278 Free PMC article. Review.
Cited by
-
Exosome-like nanoparticles from intestinal mucosal cells carry prostaglandin E2 and suppress activation of liver NKT cells.J Immunol. 2013 Apr 1;190(7):3579-89. doi: 10.4049/jimmunol.1203170. Epub 2013 Mar 6. J Immunol. 2013. PMID: 23467936 Free PMC article.
-
NF-κB is activated in CD4+ iNKT cells by sickle cell disease and mediates rapid induction of adenosine A2A receptors.PLoS One. 2013 Oct 4;8(10):e74664. doi: 10.1371/journal.pone.0074664. eCollection 2013. PLoS One. 2013. PMID: 24124453 Free PMC article.
-
Intracellular IL-4 and IFN-γ expression in iNKT cells from patients with chronic lymphocytic leukemia.Oncol Lett. 2018 Feb;15(2):1580-1590. doi: 10.3892/ol.2017.7484. Epub 2017 Nov 24. Oncol Lett. 2018. PMID: 29434853 Free PMC article.
-
NKT cell-TCR expression activates conventional T cells in vivo, but is largely dispensable for mature NKT cell biology.PLoS Biol. 2013;11(6):e1001589. doi: 10.1371/journal.pbio.1001589. Epub 2013 Jun 18. PLoS Biol. 2013. PMID: 23853545 Free PMC article.
-
Activation strategies for invariant natural killer T cells.Immunogenetics. 2016 Aug;68(8):649-63. doi: 10.1007/s00251-016-0944-8. Epub 2016 Jul 25. Immunogenetics. 2016. PMID: 27457886 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
