

Sequencing of RDR6-dependent double-stranded RNAs reveals novel features of plant siRNA biogenesis
- PMID: 22434877
- PMCID: PMC3401431
- DOI: 10.1093/nar/gks242
Sequencing of RDR6-dependent double-stranded RNAs reveals novel features of plant siRNA biogenesis
Abstract
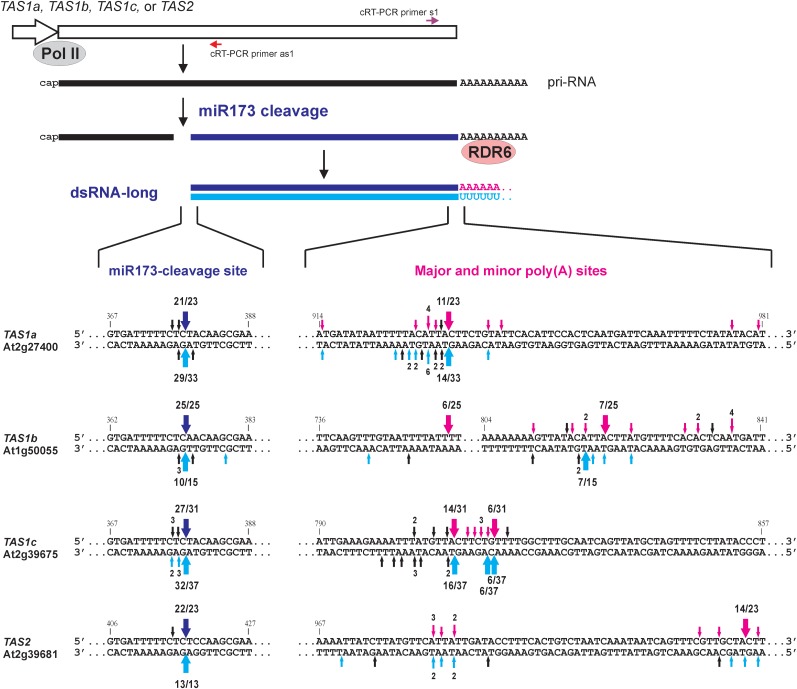
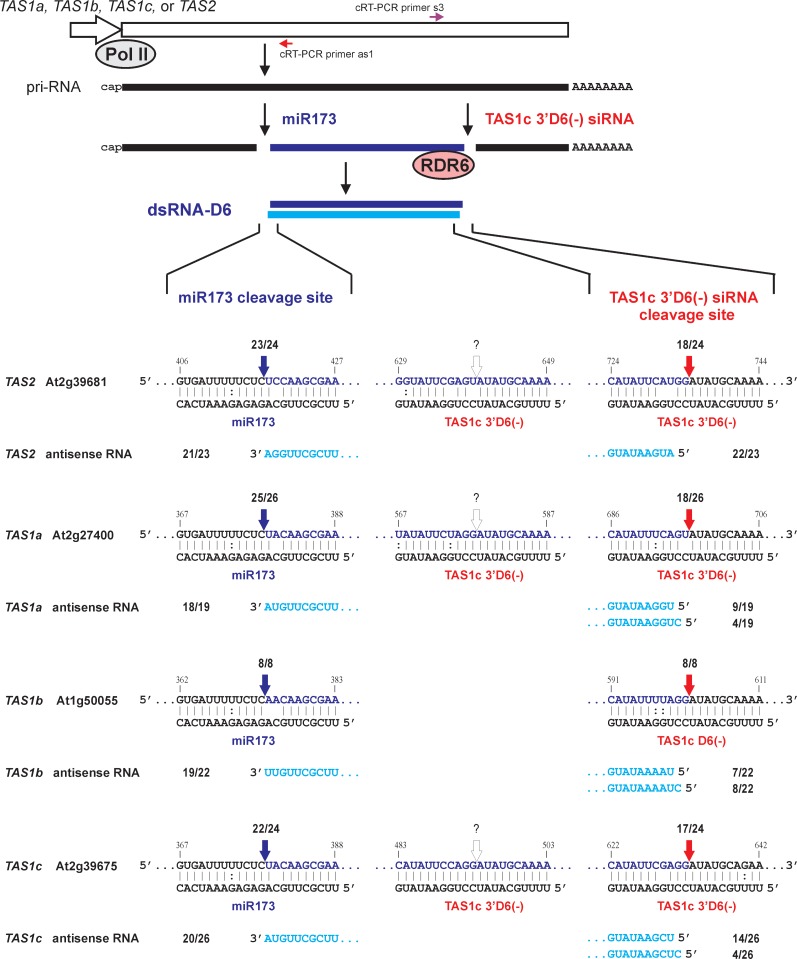
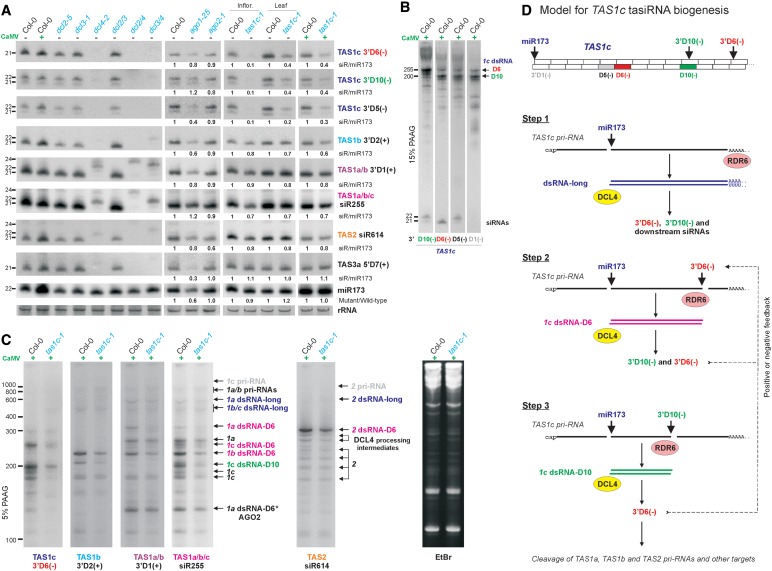
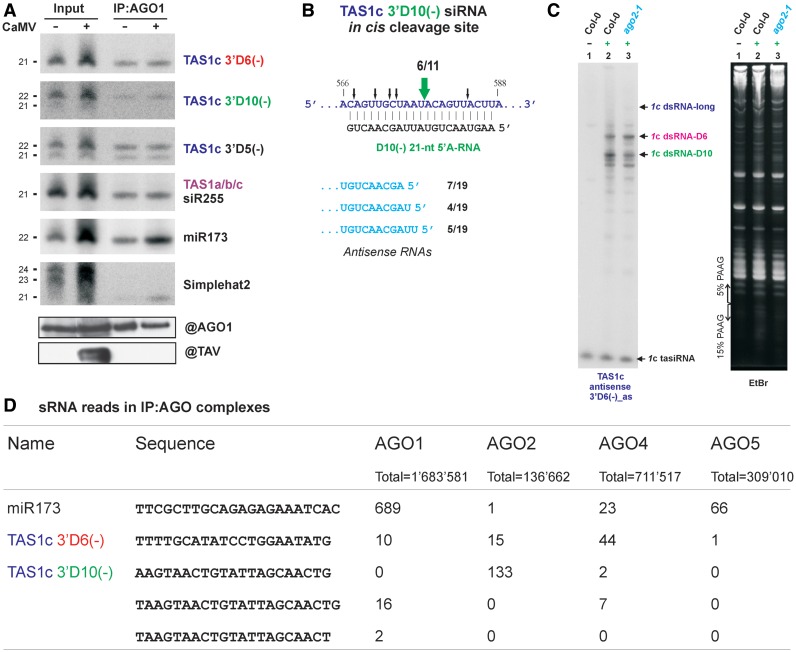
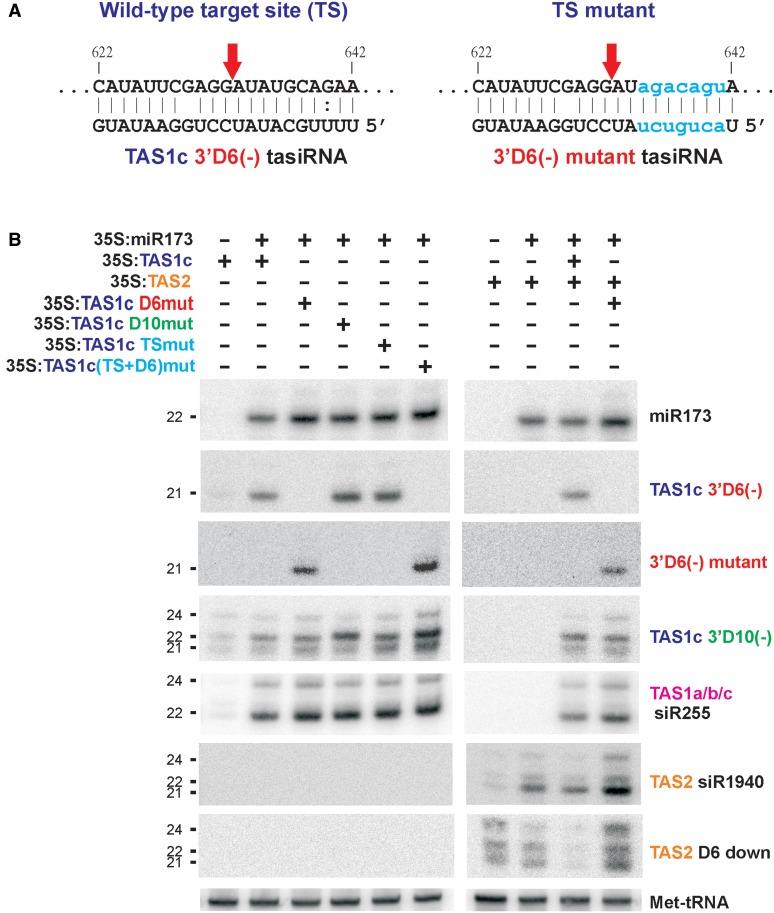
Biogenesis of trans-acting siRNAs (tasiRNAs) is initiated by miRNA-directed cleavage of TAS gene transcripts and requires RNA-dependent RNA polymerase 6 (RDR6) and Dicer-like 4 (DCL4). Here, we show that following miR173 cleavage the entire polyadenylated parts of Arabidopsis TAS1a/b/c and TAS2 transcripts are converted by RDR6 to double-stranded (ds)RNAs. Additionally, shorter dsRNAs are produced following a second cleavage directed by a TAS1c-derived siRNA. This tasiRNA and miR173 guide Argonaute 1 complexes to excise the segments from TAS2 and three TAS1 transcripts including TAS1c itself to be converted to dsRNAs, which restricts siRNA production to a region between the two cleavage sites. TAS1c is also feedback regulated by a cis-acting siRNA. We conclude that TAS1c generates a master siRNA that controls a complex network of TAS1/TAS2 siRNA biogenesis and gene regulation. TAS1/TAS2 short dsRNAs produced in this network are processed by DCL4 from both ends in distinct registers, which increases repertoires of tasiRNAs.
Figures

Similar articles
-
RDR6-mediated synthesis of complementary RNA is terminated by miRNA stably bound to template RNA.Nucleic Acids Res. 2012 Jan;40(2):594-9. doi: 10.1093/nar/gkr760. Epub 2011 Sep 19. Nucleic Acids Res. 2012. PMID: 21930511 Free PMC article.
-
Roles of dicer-like and argonaute proteins in TAS-derived small interfering RNA-triggered DNA methylation.Plant Physiol. 2012 Oct;160(2):990-9. doi: 10.1104/pp.112.200279. Epub 2012 Jul 30. Plant Physiol. 2012. PMID: 22846193 Free PMC article.
-
3' fragment of miR173-programmed RISC-cleaved RNA is protected from degradation in a complex with RISC and SGS3.Proc Natl Acad Sci U S A. 2013 Mar 5;110(10):4117-22. doi: 10.1073/pnas.1217050110. Epub 2013 Feb 15. Proc Natl Acad Sci U S A. 2013. PMID: 23417299 Free PMC article.
-
Biogenesis of trans-acting siRNAs, endogenous secondary siRNAs in plants.Genes Genet Syst. 2013;88(2):77-84. doi: 10.1266/ggs.88.77. Genes Genet Syst. 2013. PMID: 23832299 Review.
-
MicroRNA-dependent trans-acting siRNA production.Sci STKE. 2005 Sep 6;2005(300):pe43. doi: 10.1126/stke.3002005pe43. Sci STKE. 2005. PMID: 16145017 Review.
Cited by
-
An endogene-resembling transgene is resistant to DNA methylation and systemic silencing.RNA Biol. 2014;11(7):934-41. doi: 10.4161/rna.29623. Epub 2014 Jul 23. RNA Biol. 2014. PMID: 25180820 Free PMC article.
-
Investigation of a miRNA-Induced Gene Silencing Technique in Petunia Reveals Alterations in miR173 Precursor Processing and the Accumulation of Secondary siRNAs from Endogenous Genes.PLoS One. 2015 Dec 14;10(12):e0144909. doi: 10.1371/journal.pone.0144909. eCollection 2015. PLoS One. 2015. PMID: 26658695 Free PMC article.
-
Small RNAs in Plant Responses to Abiotic Stresses: Regulatory Roles and Study Methods.Int J Mol Sci. 2015 Oct 15;16(10):24532-54. doi: 10.3390/ijms161024532. Int J Mol Sci. 2015. PMID: 26501263 Free PMC article. Review.
-
Identification and molecular characterization of a trans-acting small interfering RNA producing locus regulating leaf rust responsive gene expression in wheat (Triticum aestivum L.).Planta. 2017 Nov;246(5):939-957. doi: 10.1007/s00425-017-2744-2. Epub 2017 Jul 14. Planta. 2017. PMID: 28710588
-
Coordinated Action of RTBV and RTSV Proteins Suppress Host RNA Silencing Machinery.Microorganisms. 2022 Jan 18;10(2):197. doi: 10.3390/microorganisms10020197. Microorganisms. 2022. PMID: 35208652 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
