

HIV-2 genome dimerization is required for the correct processing of Gag: a second-site reversion in matrix can restore both processes in dimerization-impaired mutant viruses
- PMID: 22419802
- PMCID: PMC3347307
- DOI: 10.1128/JVI.00124-12
HIV-2 genome dimerization is required for the correct processing of Gag: a second-site reversion in matrix can restore both processes in dimerization-impaired mutant viruses
Abstract
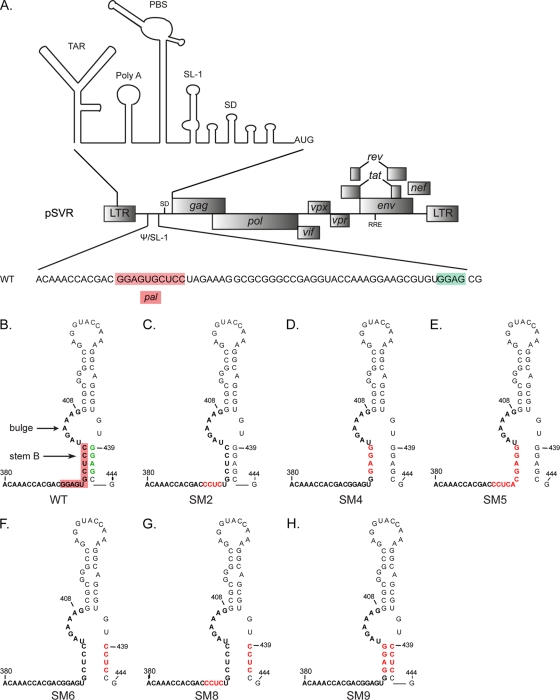
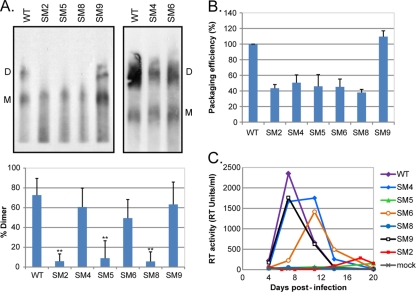
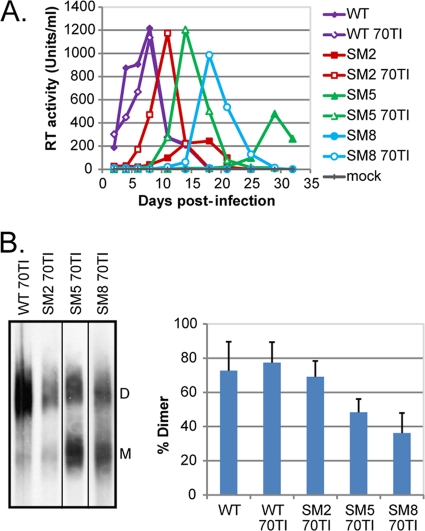
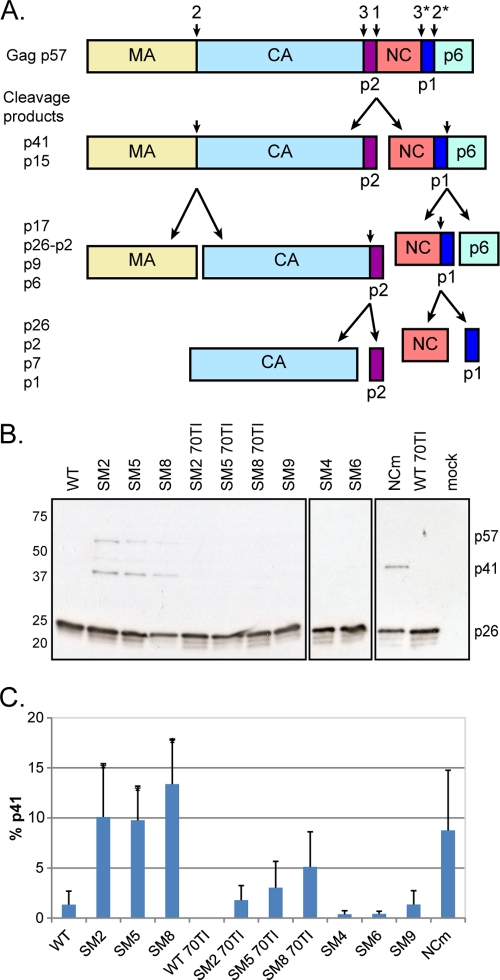
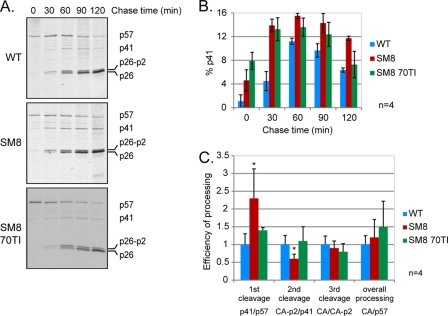
A unique feature of retroviruses is the packaging of two copies of their genome, noncovalently linked at their 5' ends. In vitro, dimerization of human immunodeficiency virus type 2 (HIV-2) RNA occurs by interaction of a self-complementary sequence exposed in the loop of stem-loop 1 (SL-1), also termed the dimer initiation site (DIS). However, in virions, HIV-2 genome dimerization does not depend on the DIS. Instead, a palindrome located within the packaging signal (Psi) is the essential motif for genome dimerization. We reported previously that a mutation within Psi decreasing genome dimerization and packaging also resulted in a reduced proportion of mature particles (A. L'Hernault, J. S. Greatorex, R. A. Crowther, and A. M. Lever, Retrovirology 4:90, 2007). In this study, we investigated further the relationship between HIV-2 genome dimerization, particle maturation, and infectivity by using a series of targeted mutations in SL-1. Our results show that disruption of a purine-rich ((392)-GGAG-(395)) motif within Psi causes a severe reduction in genome dimerization and a replication defect. Maintaining the extended SL-1 structure in combination with the (392)-GGAG-(395) motif enhanced packaging. Unlike that of HIV-1, which can replicate despite mutation of the DIS, HIV-2 replication depends critically on genome dimerization rather than just packaging efficiency. Gag processing was altered in the HIV-2 dimerization mutants, resulting in the accumulation of the MA-CA-p2 processing intermediate and suggesting a link between genome dimerization and particle assembly. Analysis of revertant SL-1 mutant viruses revealed that a compensatory mutation in matrix (70TI) could rescue viral replication and partially restore genome dimerization and Gag processing. Our results are consistent with interdependence between HIV-2 RNA dimerization and the correct proteolytic cleavage of the Gag polyprotein.
Figures
Similar articles
-
Dimerisation of HIV-2 genomic RNA is linked to efficient RNA packaging, normal particle maturation and viral infectivity.Retrovirology. 2007 Dec 13;4:90. doi: 10.1186/1742-4690-4-90. Retrovirology. 2007. PMID: 18078509 Free PMC article.
-
Compensatory point mutations in the human immunodeficiency virus type 1 Gag region that are distal from deletion mutations in the dimerization initiation site can restore viral replication.J Virol. 1998 Aug;72(8):6629-36. doi: 10.1128/JVI.72.8.6629-6636.1998. J Virol. 1998. PMID: 9658109 Free PMC article.
-
HIV-1 Matrix Trimerization-Impaired Mutants Are Rescued by Matrix Substitutions That Enhance Envelope Glycoprotein Incorporation.J Virol. 2019 Dec 12;94(1):e01526-19. doi: 10.1128/JVI.01526-19. Print 2019 Dec 12. J Virol. 2019. PMID: 31619553 Free PMC article.
-
A short sequence motif in the 5' leader of the HIV-1 genome modulates extended RNA dimer formation and virus replication.J Biol Chem. 2014 Dec 19;289(51):35061-74. doi: 10.1074/jbc.M114.621425. Epub 2014 Nov 3. J Biol Chem. 2014. PMID: 25368321 Free PMC article.
-
Structural determinants and mechanism of HIV-1 genome packaging.J Mol Biol. 2011 Jul 22;410(4):609-33. doi: 10.1016/j.jmb.2011.04.029. J Mol Biol. 2011. PMID: 21762803 Free PMC article. Review.
Cited by
-
Coordination of Genomic RNA Packaging with Viral Assembly in HIV-1.Viruses. 2016 Jul 14;8(7):192. doi: 10.3390/v8070192. Viruses. 2016. PMID: 27428992 Free PMC article. Review.
-
The matrix domain contributes to the nucleic acid chaperone activity of HIV-2 Gag.Retrovirology. 2016 Mar 17;13:18. doi: 10.1186/s12977-016-0245-1. Retrovirology. 2016. PMID: 26987314 Free PMC article.
-
Probing the Structures of Viral RNA Regulatory Elements with SHAPE and Related Methodologies.Front Microbiol. 2018 Jan 9;8:2634. doi: 10.3389/fmicb.2017.02634. eCollection 2017. Front Microbiol. 2018. PMID: 29375504 Free PMC article. Review.
-
Retroviral RNA Dimerization: From Structure to Functions.Front Microbiol. 2018 Mar 22;9:527. doi: 10.3389/fmicb.2018.00527. eCollection 2018. Front Microbiol. 2018. PMID: 29623074 Free PMC article. Review.
-
Evidence that the endosomal sorting complex required for transport-II (ESCRT-II) is required for efficient human immunodeficiency virus-1 (HIV-1) production.Retrovirology. 2015 Aug 14;12:72. doi: 10.1186/s12977-015-0197-x. Retrovirology. 2015. PMID: 26268989 Free PMC article.
References
-
- Amarasinghe GK, et al. 2000. NMR structure of the HIV-1 nucleocapsid protein bound to stem-loop SL2 of the psi-RNA packaging signal. Implications for genome recognition. J. Mol. Biol. 301:491–511 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
