

Morbillivirus glycoprotein expression induces ER stress, alters Ca2+ homeostasis and results in the release of vasostatin
- PMID: 22403712
- PMCID: PMC3293893
- DOI: 10.1371/journal.pone.0032803
Morbillivirus glycoprotein expression induces ER stress, alters Ca2+ homeostasis and results in the release of vasostatin
Abstract
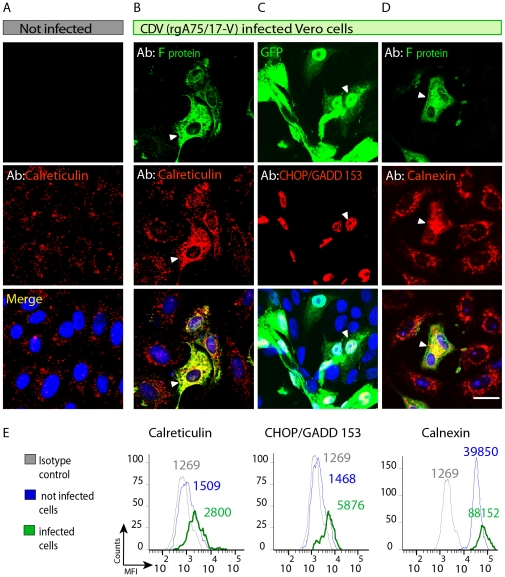
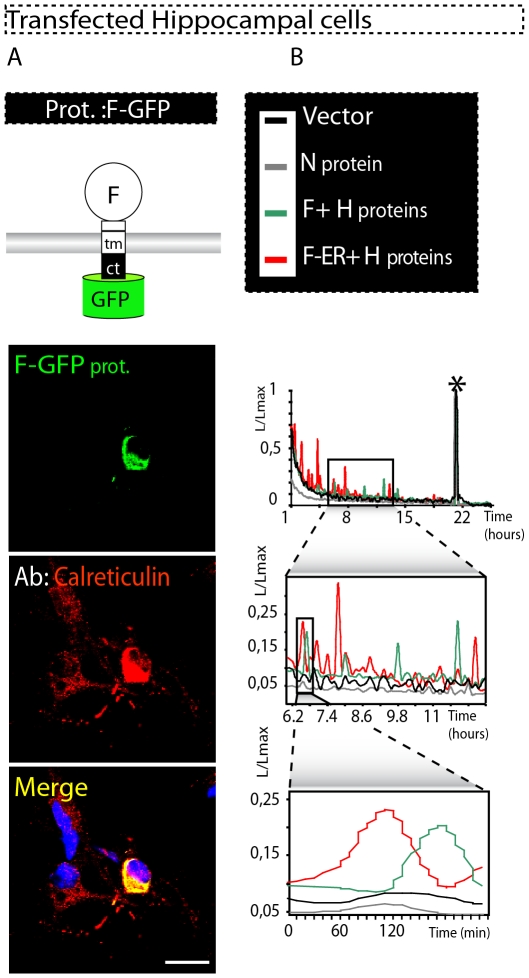
Although the pathology of Morbillivirus in the central nervous system (CNS) is well described, the molecular basis of neurodegenerative events still remains poorly understood. As a model to explore Morbillivirus-mediated CNS dysfunctions, we used canine distemper virus (CDV) that we inoculated into two different cell systems: a monkey cell line (Vero) and rat primary hippocampal neurons. Importantly, the recombinant CDV used in these studies not only efficiently infects both cell types but recapitulates the uncommon, non-cytolytic cell-to-cell spread mediated by virulent CDVs in brain of dogs. Here, we demonstrated that both CDV surface glycoproteins (F and H) markedly accumulated in the endoplasmic reticulum (ER). This accumulation triggered an ER stress, characterized by increased expression of the ER resident chaperon calnexin and the proapoptotic transcription factor CHOP/GADD 153. The expression of calreticulin (CRT), another ER resident chaperon critically involved in the response to misfolded proteins and in Ca(2+) homeostasis, was also upregulated. Transient expression of recombinant CDV F and H surface glycoproteins in Vero cells and primary hippocampal neurons further confirmed a correlation between their accumulation in the ER, CRT upregulation, ER stress and disruption of ER Ca(2+) homeostasis. Furthermore, CDV infection induced CRT fragmentation with re-localisation of a CRT amino-terminal fragment, also known as vasostatin, on the surface of infected and neighbouring non-infected cells. Altogether, these results suggest that ER stress, CRT fragmentation and re-localization on the cell surface may contribute to cytotoxic effects and ensuing cell dysfunctions triggered by Morbillivirus, a mechanism that might potentially be relevant for other neurotropic viruses.
Conflict of interest statement
Figures
Similar articles
-
SLAM- and nectin-4-independent noncytolytic spread of canine distemper virus in astrocytes.J Virol. 2015 May;89(10):5724-33. doi: 10.1128/JVI.00004-15. Epub 2015 Mar 18. J Virol. 2015. PMID: 25787275 Free PMC article.
-
Canine distemper virus infection of primary hippocampal cells induces increase in extracellular glutamate and neurodegeneration.J Neurochem. 2007 Nov;103(3):1184-95. doi: 10.1111/j.1471-4159.2007.04819.x. Epub 2007 Aug 6. J Neurochem. 2007. PMID: 17680994
-
Regulatory Role of the Morbillivirus Attachment Protein Head-to-Stalk Linker Module in Membrane Fusion Triggering.J Virol. 2018 Aug 29;92(18):e00679-18. doi: 10.1128/JVI.00679-18. Print 2018 Sep 15. J Virol. 2018. PMID: 29997204 Free PMC article.
-
Canine and Phocine Distemper Viruses: Global Spread and Genetic Basis of Jumping Species Barriers.Viruses. 2019 Oct 14;11(10):944. doi: 10.3390/v11100944. Viruses. 2019. PMID: 31615092 Free PMC article. Review.
-
Multiple Receptors Involved in Invasion and Neuropathogenicity of Canine Distemper Virus: A Review.Viruses. 2022 Jul 12;14(7):1520. doi: 10.3390/v14071520. Viruses. 2022. PMID: 35891500 Free PMC article. Review.
Cited by
-
Influenza A viruses balance ER stress with host protein synthesis shutoff.Proc Natl Acad Sci U S A. 2021 Sep 7;118(36):e2024681118. doi: 10.1073/pnas.2024681118. Proc Natl Acad Sci U S A. 2021. PMID: 34479996 Free PMC article.
-
Measles Virus Infection Fosters Dendritic Cell Motility in a 3D Environment to Enhance Transmission to Target Cells in the Respiratory Epithelium.Front Immunol. 2019 Jun 5;10:1294. doi: 10.3389/fimmu.2019.01294. eCollection 2019. Front Immunol. 2019. PMID: 31231395 Free PMC article.
-
Ebola Virus Activates IRE1α-Dependent XBP1u Splicing.Viruses. 2022 Dec 30;15(1):122. doi: 10.3390/v15010122. Viruses. 2022. PMID: 36680162 Free PMC article.
-
Marburg virus regulates the IRE1/XBP1-dependent unfolded protein response to ensure efficient viral replication.Emerg Microbes Infect. 2019;8(1):1300-1313. doi: 10.1080/22221751.2019.1659552. Emerg Microbes Infect. 2019. PMID: 31495285 Free PMC article.
-
Reduced angiogenic gene expression in morbillivirus-triggered oncolysis in a translational model for histiocytic sarcoma.J Cell Mol Med. 2017 Apr;21(4):816-830. doi: 10.1111/jcmm.13023. Epub 2016 Nov 17. J Cell Mol Med. 2017. PMID: 27860224 Free PMC article.
References
-
- Vandevelde M, Zurbriggen A. Demyelination in canine distemper virus infection: a review. Acta Neuropathol (Berl) 2005;109:56–68. - PubMed
-
- Beineke A, Puff C, Seehusen F, Baumgartner W. Pathogenesis and immunopathology of systemic and nervous canine distemper. Vet Immunol Immunopathol. 2009;127:1–18. - PubMed
-
- Brunner JM, Plattet P, Majcherczyk P, Zurbriggen A, Wittek R, et al. Canine distemper virus infection of primary hippocampal cells induces increase in extracellular glutamate and neurodegeneration. J Neurochem. 2007;103:1184–1195. - PubMed
-
- Kaufman RJ. Stress signaling from the lumen of the endoplasmic reticulum: coordination of gene transcriptional and translational controls. Genes Dev. 1999;13:1211–1233. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
