

cAMP-induced mitochondrial compartment biogenesis: role of glutathione redox state
- PMID: 22396541
- PMCID: PMC3340255
- DOI: 10.1074/jbc.M111.302786
cAMP-induced mitochondrial compartment biogenesis: role of glutathione redox state
Abstract
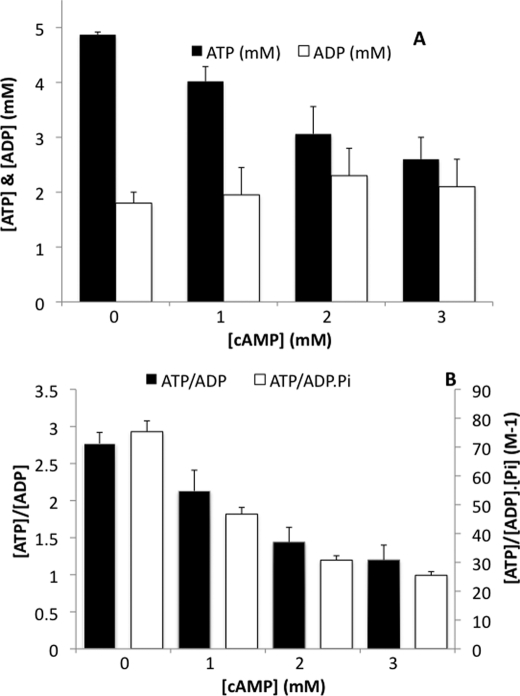
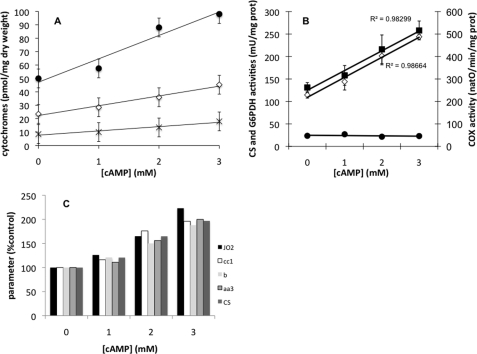
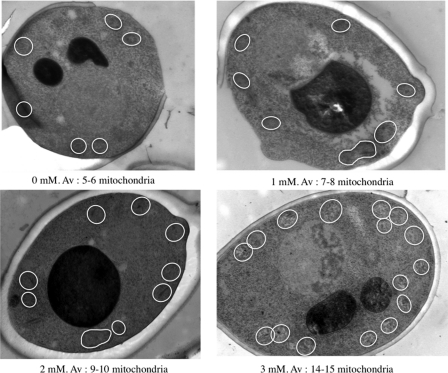
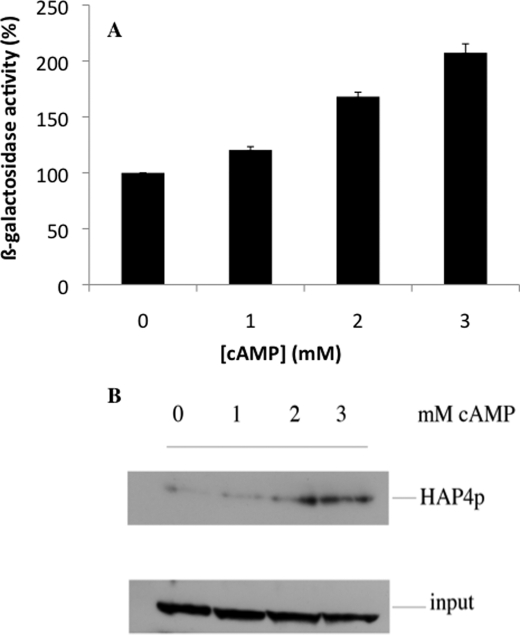
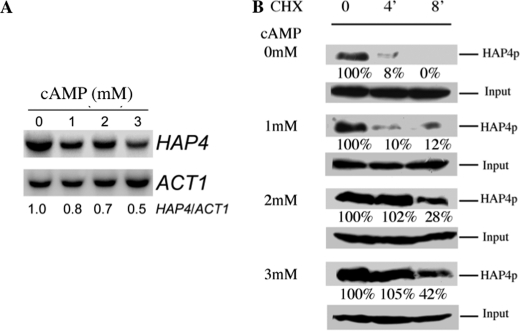
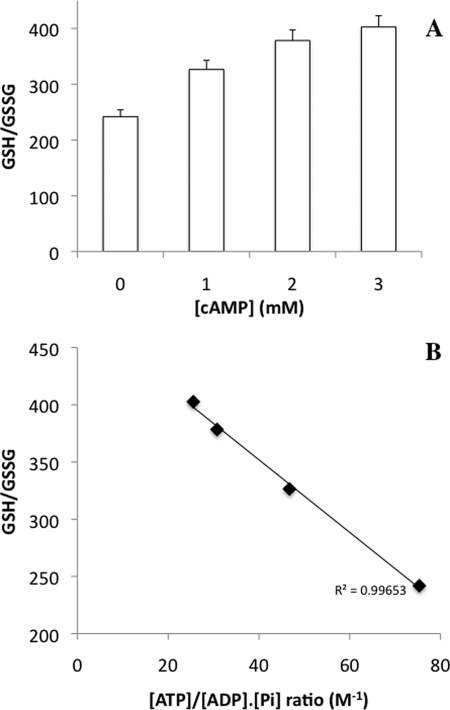
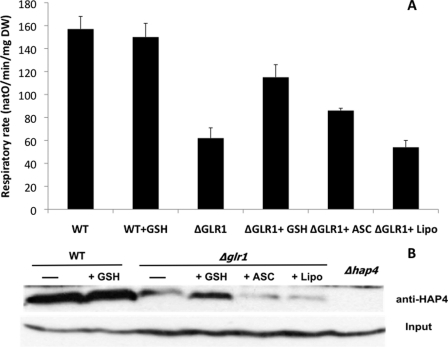
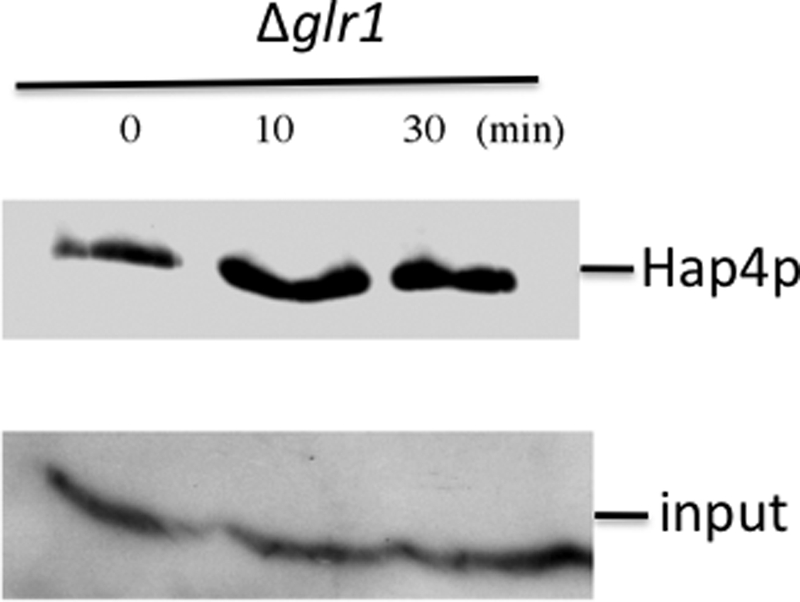
Cell fate and proliferation are tightly linked to the regulation of the mitochondrial energy metabolism. Hence, mitochondrial biogenesis regulation, a complex process that requires a tight coordination in the expression of the nuclear and mitochondrial genomes, has a major impact on cell fate and is of high importance. Here, we studied the molecular mechanisms involved in the regulation of mitochondrial biogenesis through a nutrient-sensing pathway, the Ras-cAMP pathway. Activation of this pathway induces a decrease in the cellular phosphate potential that alleviates the redox pressure on the mitochondrial respiratory chain. One of the cellular consequences of this modulation of cellular phosphate potential is an increase in the cellular glutathione redox state. The redox state of the glutathione disulfide-glutathione couple is a well known important indicator of the cellular redox environment, which is itself tightly linked to mitochondrial activity, mitochondria being the main cellular producer of reactive oxygen species. The master regulator of mitochondrial biogenesis in yeast (i.e. the transcriptional co-activator Hap4p) is positively regulated by the cellular glutathione redox state. Using a strain that is unable to modulate its glutathione redox state (Δglr1), we pinpoint a positive feedback loop between this redox state and the control of mitochondrial biogenesis. This is the first time that control of mitochondrial biogenesis through glutathione redox state has been shown.
Figures

Similar articles
-
"Labile" heme critically regulates mitochondrial biogenesis through the transcriptional co-activator Hap4p in Saccharomyces cerevisiae.J Biol Chem. 2020 Apr 10;295(15):5095-5109. doi: 10.1074/jbc.RA120.012739. Epub 2020 Feb 18. J Biol Chem. 2020. PMID: 32075909 Free PMC article.
-
Mitochondrial Biogenesis and Mitochondrial Reactive Oxygen Species (ROS): A Complex Relationship Regulated by the cAMP/PKA Signaling Pathway.Cells. 2019 Mar 27;8(4):287. doi: 10.3390/cells8040287. Cells. 2019. PMID: 30934711 Free PMC article. Review.
-
Reactive oxygen species-mediated regulation of mitochondrial biogenesis in the yeast Saccharomyces cerevisiae.J Biol Chem. 2010 Jan 15;285(3):1733-42. doi: 10.1074/jbc.M109.019570. Epub 2009 Nov 6. J Biol Chem. 2010. PMID: 19897478 Free PMC article.
-
The role of mitochondrial biogenesis and ROS in the control of energy supply in proliferating cells.Biochim Biophys Acta. 2014 Jul;1837(7):1093-8. doi: 10.1016/j.bbabio.2014.02.023. Epub 2014 Mar 3. Biochim Biophys Acta. 2014. PMID: 24602596 Review.
-
Suppression of polyglutamine-induced cytotoxicity in Saccharomyces cerevisiae by enhancement of mitochondrial biogenesis.FASEB J. 2010 May;24(5):1431-41. doi: 10.1096/fj.09-148601. Epub 2009 Dec 14. FASEB J. 2010. PMID: 20008543
Cited by
-
Reactive oxygen species-mediated control of mitochondrial biogenesis.Int J Cell Biol. 2012;2012:403870. doi: 10.1155/2012/403870. Epub 2012 May 30. Int J Cell Biol. 2012. PMID: 22693510 Free PMC article.
-
"Labile" heme critically regulates mitochondrial biogenesis through the transcriptional co-activator Hap4p in Saccharomyces cerevisiae.J Biol Chem. 2020 Apr 10;295(15):5095-5109. doi: 10.1074/jbc.RA120.012739. Epub 2020 Feb 18. J Biol Chem. 2020. PMID: 32075909 Free PMC article.
-
Mitochondrial Biogenesis and Mitochondrial Reactive Oxygen Species (ROS): A Complex Relationship Regulated by the cAMP/PKA Signaling Pathway.Cells. 2019 Mar 27;8(4):287. doi: 10.3390/cells8040287. Cells. 2019. PMID: 30934711 Free PMC article. Review.
-
Diesel exhaust particles alter mitochondrial bioenergetics and cAMP producing capacity in human bronchial epithelial cells.Front Toxicol. 2024 Jul 25;6:1412864. doi: 10.3389/ftox.2024.1412864. eCollection 2024. Front Toxicol. 2024. PMID: 39118833 Free PMC article.
-
p53 orchestrates the PGC-1α-mediated antioxidant response upon mild redox and metabolic imbalance.Antioxid Redox Signal. 2013 Feb 1;18(4):386-99. doi: 10.1089/ars.2012.4615. Epub 2012 Sep 20. Antioxid Redox Signal. 2013. PMID: 22861165 Free PMC article.
References
-
- Devin A., Rigoulet M. (2004) Regulation of mitochondrial biogenesis in eukaryotic cells. Toxicol. Mech. Methods 14, 271–279 - PubMed
-
- Devin A., Dejean L., Beauvoit B., Chevtzoff C., Avéret N., Bunoust O., Rigoulet M. (2006) Growth yield homeostasis in respiring yeast is due to a strict mitochondrial content adjustment. J. Biol. Chem. 281, 26779–26784 - PubMed
-
- Devin A., Rigoulet M. (2007) Mechanisms of mitochondrial response to variations in energy demand in eukaryotic cells. Am. J. Physiol. Cell Physiol. 292, C52–C58 - PubMed
-
- Pfeiffer T., Schuster S., Bonhoeffer S. (2001) Cooperation and competition in the evolution of ATP-producing pathways. Science 292, 504–507 - PubMed
-
- Forsburg S. L., Guarente L. (1989) Communication between mitochondria and the nucleus in regulation of cytochrome genes in the yeast Saccharomyces cerevisiae. Annu. Rev. Cell Biol. 5, 153–180 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
