

Tcra enhancer activation by inducible transcription factors downstream of pre-TCR signaling
- PMID: 22357628
- PMCID: PMC3441836
- DOI: 10.4049/jimmunol.1100271
Tcra enhancer activation by inducible transcription factors downstream of pre-TCR signaling
Abstract
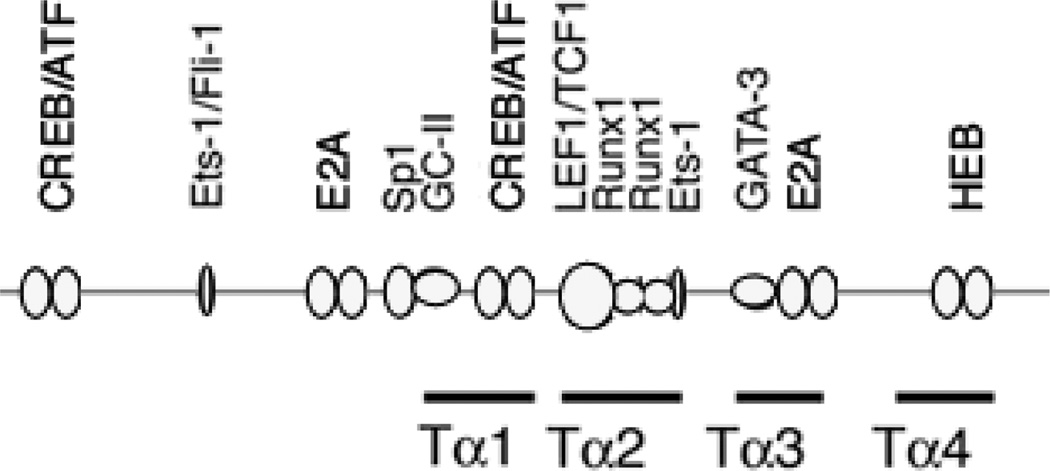
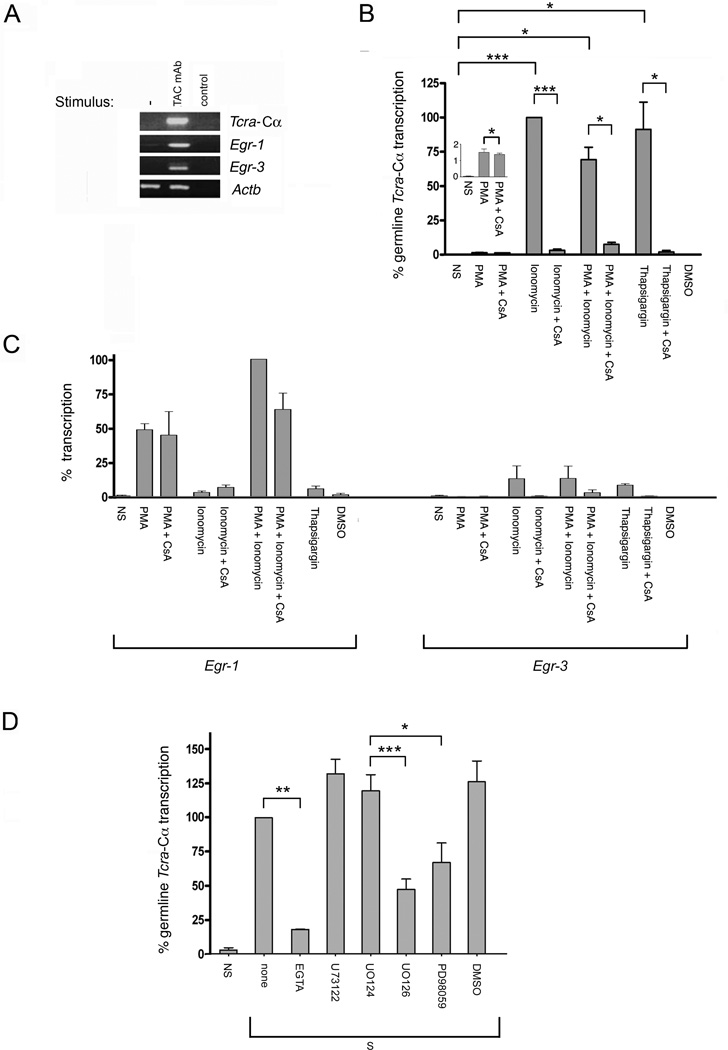
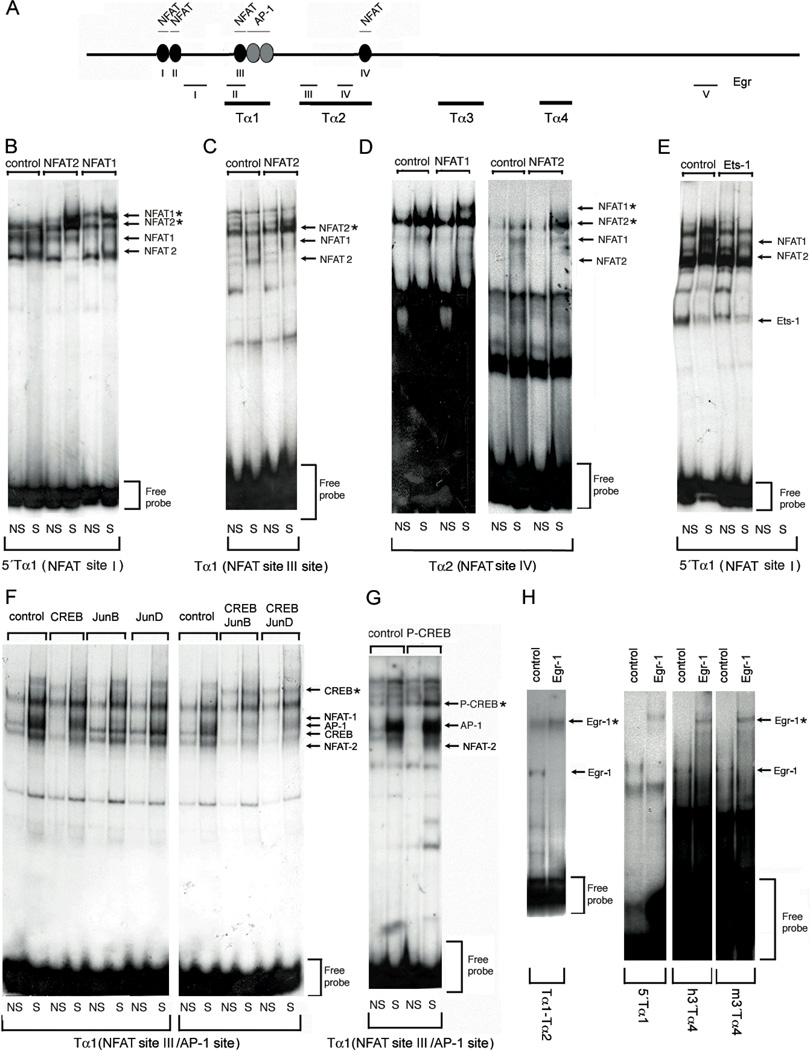
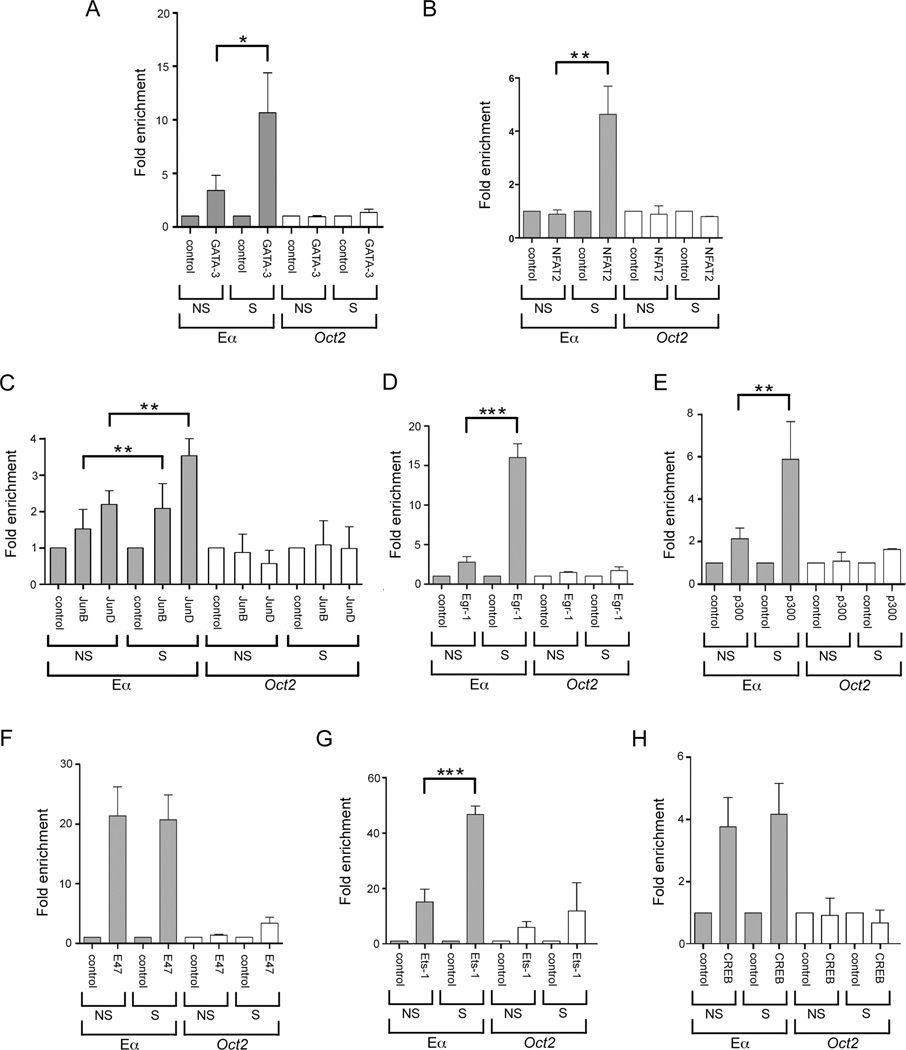
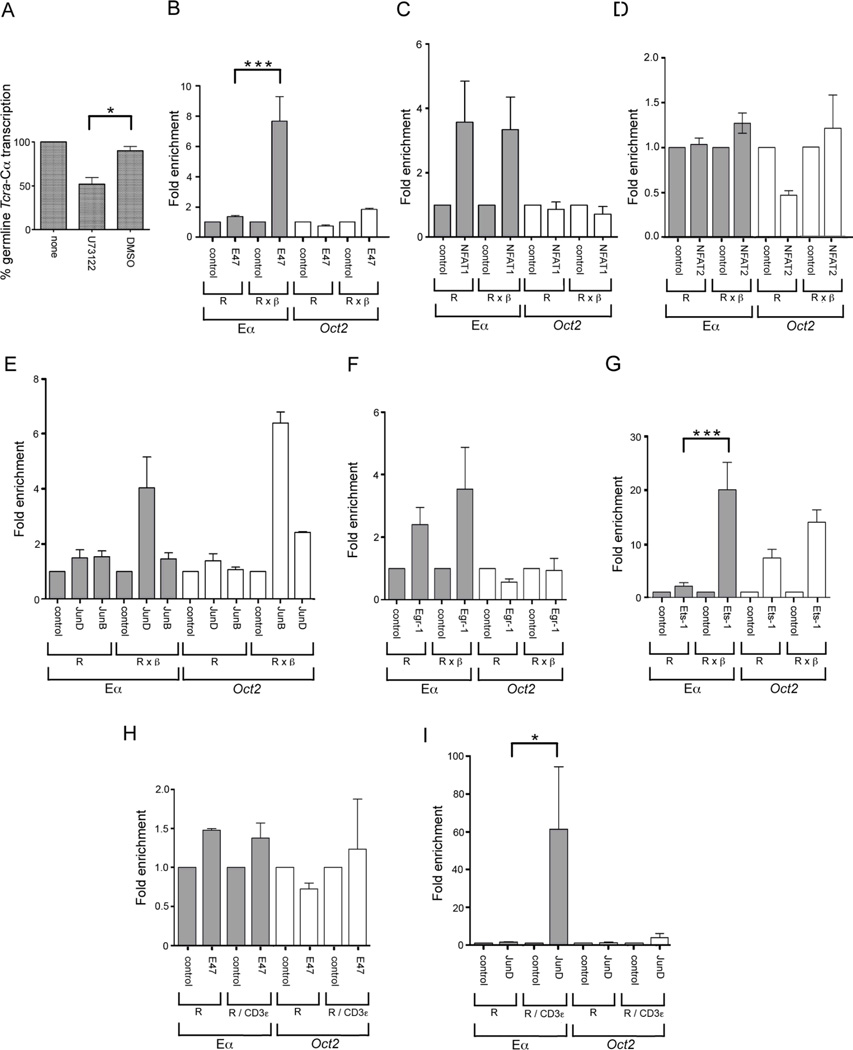
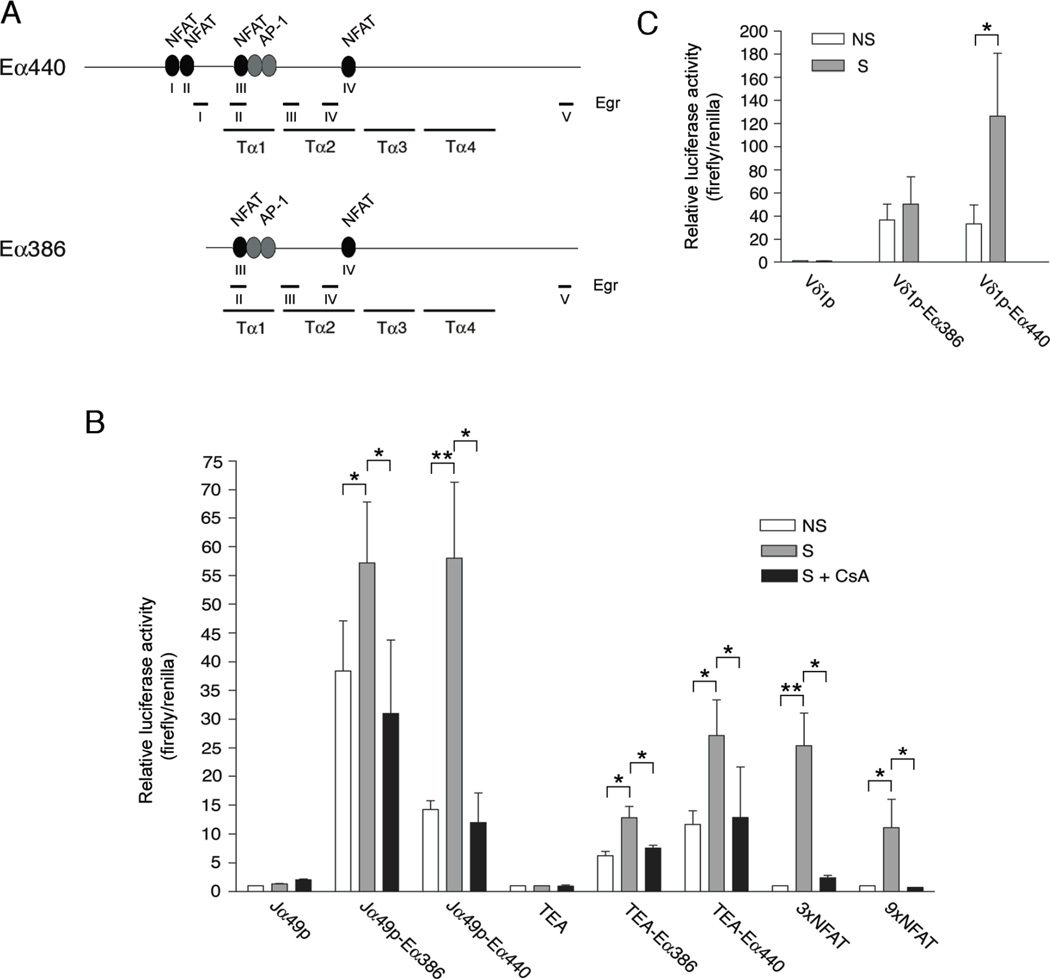
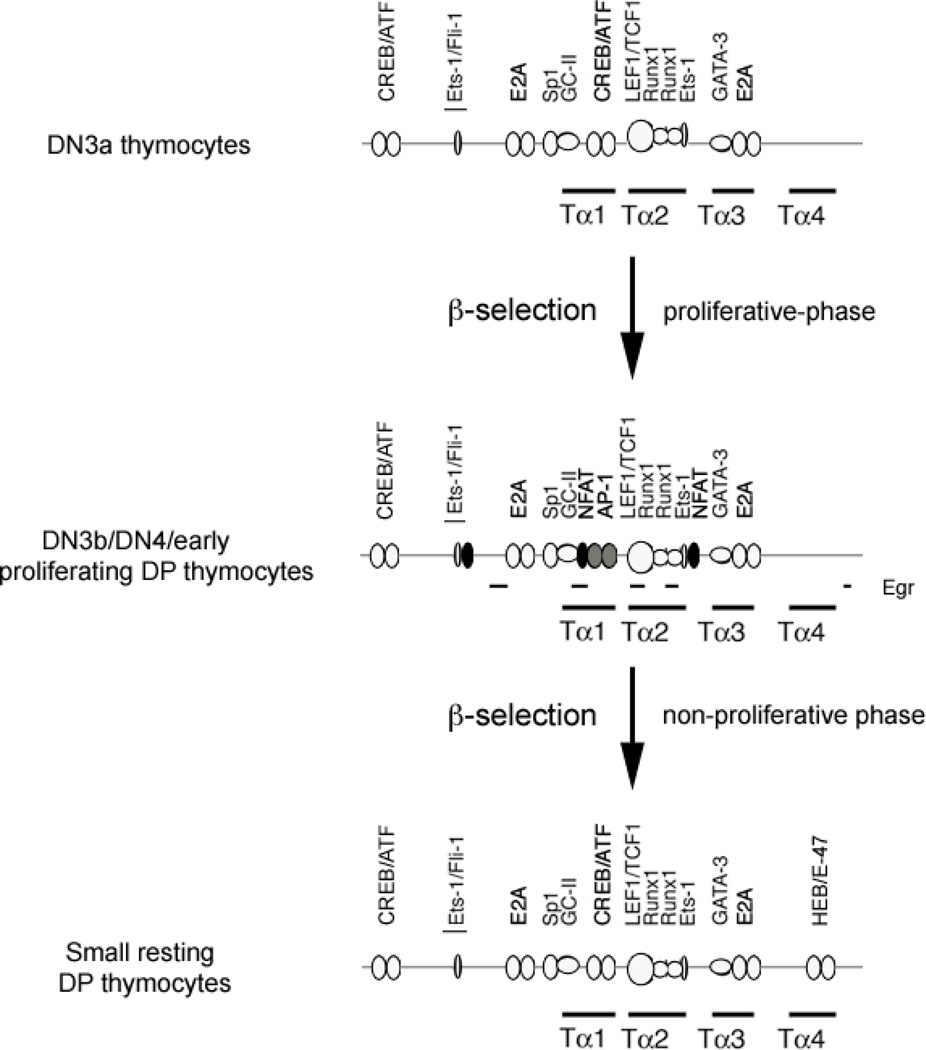
The Tcra enhancer (Eα) is essential for pre-TCR-mediated activation of germline transcription and V(D)J recombination. Eα is considered an archetypical enhanceosome that acts through the functional synergy and cooperative binding of multiple transcription factors. Based on dimethylsulfate genomic footprinting experiments, there has been a long-standing paradox regarding Eα activation in the absence of differences in enhancer occupancy. Our data provide the molecular mechanism of Eα activation and an explanation of this paradox. We found that germline transcriptional activation of Tcra is dependent on constant phospholipase Cγ, as well as calcineurin- and MAPK/ERK-mediated signaling, indicating that inducible transcription factors are crucially involved. NFAT, AP-1, and early growth response factor 1, together with CREB-binding protein/p300 coactivators, bind to Eα as part of an active enhanceosome assembled during pre-TCR signaling. We favor a scenario in which the binding of lymphoid-restricted and constitutive transcription factors to Eα prior to its activation forms a regulatory scaffold to recruit factors induced by pre-TCR signaling. Thus, the combinatorial assembly of tissue- and signal-specific transcription factors dictates the Eα function. This mechanism for enhancer activation may represent a general paradigm in tissue-restricted and stimulus-responsive gene regulation.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
T-cell receptor α enhancer is inactivated in αβ T lymphocytes.Proc Natl Acad Sci U S A. 2015 Apr 7;112(14):E1744-53. doi: 10.1073/pnas.1406551112. Epub 2015 Mar 23. Proc Natl Acad Sci U S A. 2015. PMID: 25831496 Free PMC article.
-
Recent insights into the transcriptional control of the Tcra/Tcrd locus by distant enhancers during the development of T-lymphocytes.Transcription. 2015;6(4):65-73. doi: 10.1080/21541264.2015.1078429. Transcription. 2015. PMID: 26230488 Free PMC article. Review.
-
Cooperation among multiple transcription factors is required for access to minimal T-cell receptor alpha-enhancer chromatin in vivo.Mol Cell Biol. 1998 Jun;18(6):3223-33. doi: 10.1128/MCB.18.6.3223. Mol Cell Biol. 1998. PMID: 9584163 Free PMC article.
-
Flexible stereospecific interactions and composition within nucleoprotein complexes assembled on the TCR alpha gene enhancer.J Immunol. 2009 Aug 1;183(3):1871-83. doi: 10.4049/jimmunol.0803351. Epub 2009 Jul 13. J Immunol. 2009. PMID: 19596981
-
Partners in transcription: NFAT and AP-1.Oncogene. 2001 Apr 30;20(19):2476-89. doi: 10.1038/sj.onc.1204386. Oncogene. 2001. PMID: 11402342 Review.
Cited by
-
A critical regulator of Bcl2 revealed by systematic transcript discovery of lncRNAs associated with T-cell differentiation.Sci Rep. 2019 Mar 18;9(1):4707. doi: 10.1038/s41598-019-41247-5. Sci Rep. 2019. PMID: 30886319 Free PMC article.
-
T-cell receptor α enhancer is inactivated in αβ T lymphocytes.Proc Natl Acad Sci U S A. 2015 Apr 7;112(14):E1744-53. doi: 10.1073/pnas.1406551112. Epub 2015 Mar 23. Proc Natl Acad Sci U S A. 2015. PMID: 25831496 Free PMC article.
-
Recent insights into the transcriptional control of the Tcra/Tcrd locus by distant enhancers during the development of T-lymphocytes.Transcription. 2015;6(4):65-73. doi: 10.1080/21541264.2015.1078429. Transcription. 2015. PMID: 26230488 Free PMC article. Review.
-
Interleukin-7 receptor signaling is crucial for enhancer-dependent TCRδ germline transcription mediated through STAT5 recruitment.Front Immunol. 2022 Aug 19;13:943510. doi: 10.3389/fimmu.2022.943510. eCollection 2022. Front Immunol. 2022. PMID: 36059467 Free PMC article.
-
Helix-Loop-Helix Proteins in Adaptive Immune Development.Front Immunol. 2022 May 12;13:881656. doi: 10.3389/fimmu.2022.881656. eCollection 2022. Front Immunol. 2022. PMID: 35634342 Free PMC article. Review.
References
-
- Taghon T, Yui MA, Pant R, Diamond RA, E V. Developmental and molecular characterization of emerging β- and γδ-selected pre-T cells in the adult mouse thymus. Immunity. 2006;24:53–64. - PubMed
-
- Ciofani M, Schmitt TM, Ciofani A, Michie AM, Cuburu N, Aublin A, Maryanski JL, Zúñiga-Pflucker JC. Obligatory role for cooperative signaling by pre-TCR and Notch during thymocyte differentiation. J. Immunol. 2004;172:5230–5239. - PubMed
-
- Carter JH, Lefebvre JM, Wiest DL, Tourtellote WG. Redundant role for early growth response transcriptional regulators in thymocyte differentiation and survival. J. Immunol. 2007;178:6796–6805. - PubMed
-
- Aifantis I, Gounari F, Scorrano L, Borowski C, von Boehmer H. Constitutive pre-TCR signaling promotes differentiation through Ca2+ mobilization and activation of NF-kB and NFAT. Nat. Immunol. 2001;2:403–409. - PubMed
-
- Canté-Barrett K, Winslow MM, Crabtree GB. Selective role of NFATc3 in positive selection of thymocytes. J. Immunol. 2007;179:103–110. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous